Go to John Kiernan's Facts & Files home page.
Go to
Neuroanatomy at UWO index page
Table of contents (with links to major headings)
Section 1. The nature and organization of the nervous system
Neurons and neuroglial cellsSection 2. The cells of nervous tissue: Structural aspects
Communication among cells other than neurons
Gray and white matter
Structural plan of the nervous system
The central nervous system: brain and spinal cord
The peripheral nervous system
Sensory and effector structures
NeurogliaSection 3. The cells of nervous tissue: Functional aspects
The neuron
Types of neuron
Parts of the neuron
Myelin
Nerve fibers
Synapses
Membrane potentialsSection 4. Comparative neuroanatomy
The neuronal membrane
Resting membrane potential
Depolarization and hyperpolarization
Propagation of impulses
Saltatory conduction in myelinated axons
Conduction velocity and the
compound action potential
Postsynaptic potentials: excitation
and inhibition
Neurotransmitters and neuromodulators
Axonal transport
Velocities and directions of transport
Neuroanatomical tracing methods based on
axoplasmic transport
InvertebratesSection 5. Development of the nervous system
Vertebrates
Spinal and cranial nerves
Special sense organs
Central nervous system
Why do tracts cross the midline?
Early stages in developmentSection 6. Some illustrations of animal and human neuroanatomy
Neural tube
Neural crest
Placodes
Formation of the brain and spinal cord
Histogenesis
Spinal cord, brain stem, and cerebellum
Diencephalon and telencephalon
Formation of the peripheral nervous system
Special sense organs
Developmental abnormalities of the central
nervous system
List of figures in Section 6 (with links)Section 7. A short account of the anatomy and functional pathways of the human central nervous system
Peripheral nervous system
Segmental organization
Relation of spinal cord and nerve roots to the vertebral column
Cranial nerves
Autonomic nervous system
Regional anatomy of the central nervous system
Spinal cord
Brain stem
Cerebellum
Cerebral hemisphere
Functional pathways in the central nervous system
Somatic sensations
Simple touch, temperature and pain
Discriminative touch
Proprioception
Voluntary movement: Descending motor pathways
Other circuits for movement
Cerebellar circuits
Basal ganglia circuits
Eye movements
Special senses
Equilibration
Hearing
Vision and visual reflexes
The visual pathway
Pupillary light reflex
Accommodation for near vision
Smell
Taste
Language, memory and behavior
Language and speech
Memory
Amygdala and prefrontal cortex
This web document is derived from handouts associated with Anatomy 535b (later 9535b), which was a graduate level half-course in Neuroanatomy taught in the 1990s and 2000s by me (J. A. Kiernan) in the Department of Anatomy and Cell Biology at the University of Western Ontario. That department at UWO now offers at least five courses for MSc/PhD candidates and for those in the third and fourth years of the Bachelor of Medical Sciences program, in addition to neuroanatomy contributions to the curricula for medical (MD) students and those working for master's degrees in occupational and physical therapy.
Some revisons have been made in the 13 years since I retired from teaching at UWO. This HTML document and its many links are now offered to help anyone, anywhere, wanting some quite detailed information about the nervous system.Click on links to view figures, tables or other files.
Section 1. The nature and organization of the nervous system
Multicellular animals higher on the ladder of life than the sponges all have nervous systems. A nervous system contains cells specialized for the rapid passing of signals within the animal's body. It coordinates the activity of the animal by controlling the contractile and secretary cells. The input to a nervous system comes from sensory receptors. These are cells or organs that can communicate physical or chemical events, inside or outside the body, to the cells of the nervous system. In all but the simplest animals, there are extensive connections within the nervous tissue. These encode patterns of signals that control purposeful movements, feeding, defensive and reproductive activity, and indeed the whole gamut of innate and learned behavior. Learning can occur because the intercellular circuitry of the nervous system continually adapts itself with use.
Neurons and neuroglial cells
Nervous systems range in complexity from the simple nerve net of Hydra to the mammalian nervous system consisting of brain, spinal cord, peripheral nerves, and ganglia. The conducting elements of nervous tissue are called neurons. Each neuron is a cell, usually with several long cytoplasmic processes along which the signals are carried. The nucleus and most of the synthetic machinery of the neuron are in the cell-body, or soma, from which the processes (neurites) radiate. A typical neuron has several short neurites, called dendrites (meaning little branches), that conduct principally towards the soma, and a single, longer neurite known as the axon (from the Greek for "axis"). The axon typically conducts signals away from the soma and has branches that touch the neurites of other cells. Each point of functional contact between neurons is a synapse (from the Greek verb "to join"). Nervous tissue also contains cells that do not carry signals. These are the neuroglial cells, often referred to collectively as "glia." They outnumber the neurons, with which they are intimately associated.
Communication among cells other than neurons
Nervous tissue serves the special function of communication, but there are other ways in which cells can exchange information. Gap junctions are regions of apposition of the surface membranes of cells. They occur in most embryonic tissues and in some adult tissues. Small molecules can pass freely from the cytoplasm of one cell to that of another across a gap junction. Thus gradients of concentration of developmentally significant compounds can exist across a mass of cells and may be important for growth and differentiation. Gap junctions between adjacent smooth muscle cells are important for synchronous contraction in many organs. Gap junctions also occur between certain neurons; they are called electrical synapses, and they permit coupled signaling activity of the cells. Most synapses, however, work by a chemical mechanism. Endocrine cells typically secrete hormones into the animal's circulation, and the hormones influence other cells without the necessity of close physical proximity. Non-neuronal cells may also influence each other by secreting substances into the extracellular space, but not into the general circulation. The use of short-range hormones in this way is known as paracrine secretion. Neurons often respond to circulating hormones, which thereby influence behavior. There are also neurons that release hormones into the circulation, a process termed neurosecretion.
Gray and white matter
The somata of neurons are not randomly dispersed; they are collected into aggregations of tissue known as gray matter from its color in preserved specimens. Gray matter contains cell-bodies of neurons, dendrites, and the beginning and end parts of axons. There are two main types of neuron. Principal cells have axons that leave the region and terminate in another region. Interneurons, which are smaller than principal cells, have axons that begin and end within the same region of gray matter. A circumscribed region of gray matter is a nucleus. A more or less isolated nodule containing neuronal somata is called a ganglion. (Some large nuclei in the brain are traditionally named "ganglia," though it would be simpler to confine this term to the peripheral nervous system.) Gray matter also forms extensive sheets of cortex (plural cortices) on some surfaces of the brain. White matter is white from the myelin sheaths of axons, which are the principal components of this tissue. A nerve is a thread- or cord-like bundle of axons passing among organs made of non-nervous tissue. A funiculus (from Latin, "little rope") is a major bundle of myelinated fibers in the spinal cord. A fasciculus ("little bundle") is smaller. A tract is a population of fibers en route from one region to another; often a tract occupies a distinctive fasciculus. A capsule is a conspicuous sheet of white matter in the brain. Many axons cross the midline of the body. If they connect symmetrical structures, the crossing fibers constitute a commissure. A decussation is the site at which a tract connecting asymmetric structures crosses the midline. Axons coming to a region are afferent. Axons projecting from a region are efferent (Latin prefixes for "to" or "out of," respectively, with ferre, "to carry").
Structural plan of the nervous system
In man and in all other vertebrate animals, the nervous system has two divisions: the central nervous system is contained in the axial skeleton, and the peripheral nervous system is distributed through most of the other parts of the body (Fig. 1 - Plan of generalized vertebrate nervous system).
The central nervous system: brain and spinal cord
The central nervous system is a hollow tube, the neuraxis. Its central cavity, the neurocoele, consists of the central canal of the spinal cord and the ventricular system of the brain. The structure of the neuraxis varies from place to place because of differences in the size and shape of the neurocoele and in the thickness of the wall of the tube. In higher animals, flexures of the neuraxis help to fit the essentially tubular brain into a round head. One end of the tube points towards the nose or beak (rostrum) and the other points towards the tail (cauda). Relative positions along the length of the central nervous system are defined by the terms rostral and caudal. These words are less ambiguous than the general (human) anatomical terms "superior," "inferior," "anterior," and "posterior." The ventral surface of the neuraxis faces the alimentary canal and the other internal organs; the dorsal surface faces in the opposite direction, towards the back and the top of the head. The midline is the plane that bisects the body, including
the nervous system, into the left and right halves. Nearness to the midline is indicated by the word medial, whereas lateral denotes remoteness from the midline. A median structure is within, or astride, the midline.
The rostral end of the central nervous system, contained in the cranium, is the brain. The caudal end of the brain continues into the spinal cord. This is contained in the spinal canal formed by the vertebrae and their connecting ligaments. The cranial cavity becomes continuous with the spinal canal through the foramen magnum, a large hole in the occipital bone. A foramen (from the Latin forare, to pierce) is a hole. Nerves connected with the brain or spinal cord pass through foramina in the base of the skull or between the vertebrae, respectively.
The spinal cord, like the vertebral column, is segmented, though the segments blend imperceptibly into each other. Left and right spinal nerves, one pair per segment, pass through the intervertebral foramina and are distributed to the trunk, appendages, and viscera. These nerves belong to the peripheral nervous system. There is an abrupt transition between central and peripheral nervous tissue at the surface of the cord. Equivalent junctions exist where the cranial nerves connect with the brain.
The brain also develops from segments known as neuromeres, but this fact cannot be discerned by simple inspection. The brain is formed of three main parts, hindbrain, the midbrain, and the forebrain. The hindbrain, which merges caudally with the spinal cord, consists of the medulla oblongata (usually called simply the medulla) caudally, and the pons rostrally. The midbrain (or mesencephalon) consists of the tectum dorsally and the two cerebral peduncles ventrally. The forebrain is the most rostral part of the brain. The caudal part of the forebrain is the diencephalon; the rostral part is the telencephalon. (These rather cumbersome terms are made up from Greek roots that mean "between-brain" and "end-brain," respectively.)
Dilatations of the neurocoele within the brain are called ventricles. The hindbrain contains the fourth ventricle. This is continuous caudally with the central canal of the spinal cord and rostrally with the cerebral aqueduct, which is the cavity of the midbrain. In the diencephalon the cavity becomes the third ventricle. Further rostrally the neurocoele bifurcates, so that the third ventricle leads into left and right lateral ventricles. Thus, the telencephalon consists of two cerebral hemispheres, each containing a lateral ventricle.
The neurocoele contains cerebrospinal fluid. This is secreted by choroid plexus, a vascular tissue that intrudes into the ventricles.
The shape of the brain is due partly to the ventricular system and partly to the variable thickness of the nervous tissue forming its walls. Some conspicuous and important parts of the central nervous system do not have central cavities continuous with the neurocoele, at least in adult life. The cerebellum is a large outgrowth of the dorsal and lateral surfaces of the hindbrain and midbrain. The optic nerve and the retina of the eye are outgrowths from the diencephalon. The diencephalon also has two glandular outgrowths: the epiphysis (or pineal gland) dorsally, and the hypophysis (or pituitary gland) ventrally. The olfactory bulbs are paired, stalked structures in the rostral part of the telencephalon. They are concerned with the sense of smell. The most conspicuous part of the a mammalian brain is the cerebral cortex, which covers the surfaces of the cerebral hemispheres (Fig. 2 - Whole human CNS and some nerves).
The peripheral nervous system
The central nervous system is connected to other parts of the body by nerves. Neuronal cell-bodies outside the central nervous system occur in ganglia (singular, ganglion). The spinal nerves are segmentally organized. Each has a dorsal and a ventral root, separately connected with the spinal cord. The dorsal root bears a ganglion (called either a spinal ganglion or a dorsal root ganglion); the ventral root does not. The dorsal roots are exclusively sensory in mammals, whereas the ventral roots contain the axons of motor neurons and the axons of neurons that control internal organs, blood vessels, and glands.
Paired cranial nerves connect the brain with other structures. The olfactory nerves, concerned with smell, enter the olfactory bulb, which is at the rostral end of the telencephalon. The optic nerves, like the retinas of the eyes, are made of central nervous tissue. They are therefore not real nerves but outgrowths of the brain. The remaining cranial nerves emerge from the brain stem, which consists of the midbrain pons and medulla.
Sensory and effector structures
The word receptor has two different meanings in neurobiology:
1. A macromolecule (on the surface of a cell, or sometimes inside a cell), which selectively binds molecules that initiate a response in the cell. Drugs, hormones, and the transmitter substances used for communication among neurons act upon cells by combining with receptor molecules.
2. A structure that acts as a sense organ, mediating conduction of signals into the nervous system.
An effector is an arrangement of contractile or secretary cells that either moves or secretes a product in response to activity in its afferent nerve.
Sensory Receptors. The simplest sensory receptors are the terminal branches of the axons of primary sensory neurons. The cell-bodies are in the ganglia of the dorsal spinal nerve roots and of cranial nerves. Free nerve endings are widely distributed in skin, blood vessels, viscera, connective tissue, and joints. Occasionally, as in hair follicles, there may be an ordered arrangement of the axons, but usually there is not obvious organization. Receptors of this kind respond to a wide variety of physical and chemical stimuli. Individually, however, they are specialized for particular sensations such as pain, temperature change, mechanical deformation, and the detection of substances in the extracellular fluid. The stimulus directly excites the surface membrane of the axon.
In structurally more complicated receptors, the axonal endings are associated with special cells that form a capsule, giving the sense organ a distinctive appearance under the microscope. These receptors usually detect noninjurious mechanical stimuli. The cells of the capsule may be involved in the process of transduction, which is the conversion of one form of energy into another. All receptors and effectors are transducers, and some can be validly compared to such inanimate electrical devices as the microphone, the television camera, and the electric motor.
Special receptors exist that generate neural signals from light, sound, position and movement of the head, and chemical stimuli in the mouth and nose. These sense organs all contain cells specialized for transduction, together with organized frameworks of supporting cells.
Movement and secretion. The most conspicuous effector cells are skeletal striated muscle fibers, each consisting of a cytoplasmic tube containing many nuclei and a highly organized system of contractile filaments. The fibers are collected together to form muscles, which make up much of the bulk of the body. The ends of most muscles are attached, often through collagenous tendons, to the skeleton. Each skeletal muscle fiber is contacted by a branch of the axon of a motor neuron, whose cell-body is in either the brain stem or the spinal cord. A train of impulses in the motor axon stimulates the release of acetylcholine at the neuromuscular junction. The acetylcholine acts only briefly, because its destruction is catalyzed by an enzyme, acetylcholinesterase. Released acetylcholine triggers a series of changes in the muscle fiber leading to contraction. The coordination of the contractions of muscle fibers within different muscles results in purposeful movement. This coordination is a major function of the central nervous system.
The effector tissues in viscera are smooth muscle, cardiac muscle (only in the heart), and the secretory cells of glands. Some glands also contain contractile myoepithelial cells that serve to squeeze the secreted products into the ducts. Visceral effector cells are innervated by neurons whose cell-bodies lie in ganglia. The innervating neurons are themselves contacted by the axons of neurons whose somata are in the brain stem or spinal cord. Thus, a chain of at least two neurons is involved in the control of any visceral structure by the central nervous system (Fig. 3 - Somatic and visceral innervation).
Section 2. The cells of nervous tissue: Structural aspects
This section and the next are concerned with the cell biology of normal nervous tissue. The constituent cells are described, and then the electrical and chemical mechanisms of signaling are explained. Most of the information in these two sections applies to all nervous systems, including that of man. Much of the knowledge of neuronal function has been obtained from the nervous tissues of submammalian vertebrates and invertebrates.
The nervous system contains neurons, which are specialized for communication, and neuroglial cells (or gliocytes), which provide structural and metabolic support for the neurons. There is very little connective tissue in the central nervous system. Much more is present in the peripheral nervous system, because the nerves and ganglia are not protected by enclosure in the axial skeleton.
The central nervous system develops from an embryonic structure, the neural tube, and it remains hollow when fully grown. The peripheral nervous system develops from the paired neural crests, which are populations of cells that lie alongside the neural tube. All the neurons and gliocytes of the central nervous system are descendents of the cells that line the lumen (neurocoele) of the neural tube. Nearly all the neurons and gliocytes are produced before birth, but some neuroglial cells continue to be produced in adult animals, principally in a "subventricular zone" close to the lining of the neurocoele. This region also contains some stem cells that are potentially able to generate new neurons. The neural crest gives rise to the neurons and neuroglia of the peripheral nervous system.
Neuroglia
The word neuroglia means "nerve-glue" (from two Greek words); it is often shortened to glia. When the term was coined, it was thought that the tissue consisted of extracellular fibrillary and granular material in addition to cells. It is now known that the neuroglia consists entirely of cells, and that the fibrous and particulate elements are cytoplasmic organelles. The different types of neuroglial cells are described in Table 1 - Neuroglia details and illustrated in Fig. 4 - Types of glial cells.
Click on highlighted links to view figures or tables.
The neuron
A neuron is a cell whose main function is rapid intercellular communication. Its characteristic features are a soma (or cell body) with long cytoplasmic processes (neurites = dendrites + axon) and points of functional intercellular contact (synapses). Rapid signaling occurs by means of electrical changes in the surface membrane of the neuron. Slower functions are mediated by the transport of many substances within the axon, both to and from the cell-body.
Types of neuron
The shapes of some typical neurons are shown in Fig. 5 - Shapes and sizes of neurons. There is great variety of shape and size, length of axon, and richness of branching of dendrites. Each type of neuron, however, is fairly constant in its shape, and each type is found in its own anatomical site within the nervous system. It should be noted that in most types of neuron the dendrites receive signals and the axon conducts them away from the region of the soma.
Generally speaking, there are two types of neuron: a principal cell has a large somata and a long axon that connect one region of the nervous system with another; an interneuron has a small soma, and its short axon is confined to the region in which the cell occurs. (Some interneurons have no axon.)
Parts of the neuron
The part of the neuron most important for the conduction and transmission of signals is the surface membrane, or plasmalemma.
The nucleus of a large neuron is large and empty-looking. The chromatin is visible only as a darkly staining or electron-dense rim within the nuclear membrane, but there is a prominent nucleolus. Small neurons often have dense nuclei, similar to those of oligodendrocytes. The cytoplasm of the soma is dominated by the organelles of protein synthesis (rough endoplasmic reticulum and polyribosomes) and cellular respiration (mitochondria). There is also a well developed Golgi apparatus, where carbohydrate side-chains are added to protein molecules destined to enter or pass through the surface membrane of the cell. In light microscopy the rough endoplasmic reticulum is conspicuous as bodies of Nissl substance, named after Franz Nissl (1860-1919), a German psychiatrist.
Fibrous organelles are the most conspicuous components of the neurites. Microfilaments 4-5 nm in diameter, made of actin, occur on the inner surface of the cell membrane. They are also abundant in growth cones, which are the motile expanded tips of growing neurites. Intermediate filaments (9-10 nm) known as neurofilaments are most abundant in axons. Microtubules (24-25 nm external diameter) occur in all parts of the neuron; they are most conspicuous in dendrites and at the axonal hillock. The latter is the site on the soma, or on a large dendrite, from which the axon arises; it is also called the "initial segment" of the axon. Neurites also contain mitochondria and fragments of smooth endoplasmic reticulum.
Dendrites taper with distance from the soma, and the diameters of successive branches become smaller. Synapses (to be described later) are present over most of the surface of a dendrite. An axon has the same diameter along its whole length, and forms synapses in its preterminal and terminal parts. The axonal cytoplasm is called axoplasm, and the surface membrane is called the axolemma.
Myelin
Many axons are ensheathed in myelin. This is a close wrapping of many layers of double membrane derived from the ensheathing glial cell. The myelin sheath is formed by elongation and rolling of the mesaxon (Fig. 6 - Ensheathment of axons). The roll is so tight that the cytoplasm and extracellular fluid are squeezed out from between the layers of membrane. A Schwann cell myelinates only one axon, but in the central nervous system each process of a single oligodendrocyte contributes to the myelination of a different axon. (Theodor Schwann (1810-1882), the German anatomist who described the neurolemmal cell, was also an originator of the "cell theory," which maintained that all organisms were composed of separate, living cells.)
The segment of an axon myelinated by a single Schwann cell or oligodendrocyte process is called an internode. Between the internodes are short interruptions (nodes of Ranvier) in the continuity of the sheath. (Louis Antoine Ranvier (1835-1922) was a French histologist and physician.) At these points the surface membrane of the axon, the axolemma, is in contact with the extracellular fluid. The electrical events of conduction occur at the nodes of a myelinated axon, and this provides for faster conduction of impulses than would be possible in an unmyelinated axon. Accordingly, myelinated axons occur in peripheral nerves and in the long tracts of the central nervous system. The unmyelinated axons in nerves are involved in functions for which great speed is not essential: some types of pain, and the innervation of blood vessels, glands, and internal organs.
Nerve fibers
A nerve fiber is an axon together with myelin sheath, if present, and the ensheathing glial cells. The velocity of conduction of an impulse along a nerve fiber increases with the diameter. The largest axons have the thickest myelin sheaths and, therefore, the greatest external diameters. The axonal diameter is approximately two thirds of the total external diameter of the fiber. The thinnest, most slowly conducting axons are unmyelinated.
In some invertebrates, notably annelids and cephalopods, there are unmyelinated axons 1 mm or more in diameter, known as giant fibers. These provide rapid conduction in animals that cannot form myelin, and they are involved in the control of movements that enable these creatures to escape from danger. Cyclostomes (lampreys and hagfish) have no myelinated axons, but in their spinal cords are the large (50 æm) unmyelinated axons of the Muller cells, whose somata are in the hindbrain. All other vertebrates have myelinated axons for rapid conduction, so there is no need for unmyelinated fibers more than about 3 æm in diameter. Mauthner cells occur in the hindbrains of fishes and larval amphibians, one cell only on each side of the midline. Each of these huge neurons has a large axon that crosses the midline and then goes caudally in the spinal cord, to the neurons supplying the muscles of the tail. The Mauthner fibers are large, and in adult fishes they are myelinated. They are for stimulating fast, powerful movements used in escaping.
Peripheral nerve fibers are classified into groups according to external diameter and conduction velocity (Table 2 - Nerve fiber types). Axons in the central nervous system are not as easy to classify; their diameters vary greatly.
Synapses
A point of functional contact between two neurons, or between a neuron and an effector cell, is a synapse. The structural details of synapses can be resolved only by electron microscopy. Most synapses in vertebrate animals are chemical synapses. The surface membranes of the two cells are thickened by deposition of fibrillary material on the cytoplasmic sides. The intervening gap contains an electron-dense glycoprotein that is absent from the general extracellular space.
The presynaptic neurite, which is most often a branch of an axon, is known as a synaptic terminal or bouton terminal ("terminal button"; the plural is boutons terminaux). This old term recalls the appearance in light microscopy.) A synaptic terminal contains numerous mitochondria and a cluster of synaptic vesicles. The latter are membrane-bound organelles 40 to 150 nm in diameter. According to the type of synapse, the vesicles may be spherical or ellipsoidal, and they may or may not have electron-dense cores. More than one type of vesicle may be present in a single terminal. Synaptic vesicles contain the chemical neurotransmitters that are released into the synaptic cleft to act upon the postsynaptic membrane.
The postsynaptic structure is typically a dendrite. Often it bears a pendunculated projection, a dendritic spine, that invaginates the presynaptic neurite. Commonly, synapses are grouped together on a dendrite or an axonal terminal to form a larger structure, known as synaptic complex or glomerulus. In the central nervous system, the cytoplasmic processes of protoplasmic astrocytes intimately invest synaptic complexes, probably to restrict diffusion in the intercellular spaces of transmitters and inorganic ions such as calcium and potassium.
Some different types of chemical synapse are shown in Fig. 7 - Various synapses. The most common arrangements for transferring signals from one neuron to another are axodendritic and axosomatic synapses. Axoaxonal synapses are strategically placed to interfere either with the initiation of impulses at the initial segments of other axons, or with the activities of other synaptic terminals. Dendrodendritic synapses can modify a neuron's responses to other synapses.
The other type of functional contact between neurons is the electrical synapse. The cell membranes, usually of neuronal somata, are closely approximated (3 nm, in contrast to 25 nm for a chemical synapse). Large, tubular protein molecules bridge the cleft and are embedded in the surface membranes of both cells, providing channels through which inorganic ions, water, and other small molecules can pass. Thus an electrical change in one of the neurons is immediately propagated to the other, and the two cells are electrotonically coupled. The electrical synapse is identical to the gap junction or nexus found in many non-nervous tissues.
Section 3. The cells of nervous tissue: Functional aspects
Depolarization and hyperpolarization
In the dendrites and soma of a neuron, the changes in membrane potential are graded; they vary in time and space with the incoming synaptic activity. The axon, on the other hand, conducts impulses, which are waves of complete depolarization of the membrane, in an all-or-none fashion. Conduction in unmyelinated axons will be considered first. The physics and chemistry of conduction were discovered in the giant axons of the squid (see Table 2), which are easier to work with than the nerve fibers of vertebrates. The mechanisms are now known to be similar in all animals.
Consider a point on an axon before, during, and after the passage of an action potential. The following events occur.
1. The imminent arrival of the impulse causes a reduction of the membrane potential from about -70 to about -20 mV.
2. This amount of reduction in potential is the threshold for depolarization, and it opens the voltage-gated sodium channels. Sodium ions immediately move into the axoplasm. They do this because (a) they are attracted by the negative charge inside, and (b) the concentration of Na+ outside is much higher than inside. The inrush of Na+ causes reversal of the membrane potential to about +40 mV in less than 1 millisecond.
3. The sodium channels close, and the influx of Na+ ceases. This is called inactivation of the channels. At the same time, the voltage-gated potassium channels open in response to the depolarization caused by the incoming Na+. Potassium ions now move out of the axon. They do so because (a) they are, at this moment, electrically repelled by the intra-axonal excess of positive charge due to the influx of Na+, and (b) the concentration of K+ inside is much higher than outside.
4. The outward diffusion of K+ takes about 2 milliseconds to restore the membrane potential to its original -70 mV. The recovery is assisted by the sodium pumps, which expel Na+ and pull in K+. The membrane becomes slightly hyperpolarized (-80 mV) at this time, for about 1 millisecond.
5. While the membrane potential is being restored to the resting level of -70 mV, the sodium channels remain closed (inactivated), so the membrane cannot be depolarized. It is said to be refractory. The axon is refractory for about 2 milliseconds after the passage of an action potential; this prevents backward propagation of the impulse.
6. The depolarization due to influx of Na+ spreads in both directions, lowering the membrane potential to the threshold value of about -20 mV. This change has no effect on the refractory membrane that is still recovering from the passage of the action potential. In the forward direction, however, the axonal membrane is not refractory, so reduction of the potential to -20 mV opens the gates of the sodium channels. There is a rapid influx of Na+, and the membrane is depolarized.
7. The continuous repetition of this cycle of ionic movements results in propagation of the action potential in one direction.
The events are shown graphically in Fig. 8 - Action potential propagation.
The ionic movements and resultant electrical changes are not the same in all neurons. Gated channels exist for ions other than Na+ and K+. Calcium channels have special significance in the presynaptic parts of axons. In the resting state, the concentration of Ca2+ is always much lower in cytoplasm than in extracellular fluid, and most of the cytoplasmic Ca2+ is sequestered by calcium binding proteins. When an action potential arrives, calcium channels open, and at presynaptic sites the influx of free Ca2+ ions triggers the secretion of neurotransmitter molecules into the synaptic cleft. Calcium ions, like sodium ions, are removed from cytoplasm by an energy-consuming pumping mechanism.
Saltatory conduction in myelinated axons
The velocity of conduction of an action potential along an unmyelinated axon increases in proportion to the square root of the diameter. Thus a mammalian unmyelinated fiber (0.2-1.5 æm) conducts at 0.5 to 2.5 meters per second, and a squid's giant axon (1.0 mm) conducts at about 25 meters per second. An advanced nervous system, which needs great numbers of rapidly conducting axons, would be impracticably large if it had to rely on unmyelinated nerve fibers.
Myelination allows high conduction velocities (up to 120 meters per second) without an inordinate increase in diameter. In a myelinated axon, all the sodium and potassium channels are concentrated at the nodes. The internodes are electrically insulated by the layers of membrane that make up the myelin sheath. The myelin accounts for about one third of the total diameter of a nerve fiber, and the length of an internode is about 100 times the external diameter.
The ionic movements of an action potential can occur only at the nodes, but electrical conduction along the internodal axon, which behaves as a well-insulated wire, reduces the membrane potential to its threshold level at the next node. Thus the impulse jumps quickly from node to node. This form of propagation is called saltatory conduction (from the Latin saltare, to jump).
Conduction velocity and the compound action potential
The fibers in mammalian peripheral nerves are classified as in Table 2. Comparable populations of axons exist in the central nervous system, but are not included in any generally recognized system of classification.
In the names of fiber-types in the peripheral nervous system, the letters A, B, and C (and the subtypes alpha, beta, gamma and delta of Group A) come from the phases of the compound action potential. This is a response recorded by an electrode in contact with a whole nerve. Following a brief electric shock at a distant point on the nerve, action potentials are initiated and propagated in all the axons (Fig. 9). These impulses reach the recording electrode at different times, determined by the conduction velocities of the axons. It is possible to dissect successively thinner strands from a nerve or a nerve root until one is obtained that contains only a single functioning nerve fiber. The action potentials recorded from such individual fibers can be related to function. The Roman numerals used to name sensory fibers were originally used in studies of single fibers dissected from dorsal spinal roots.
Postsynaptic potentials: excitation and inhibition
At a chemical synapse, the arrival of an impulse at the presynaptic terminal depolarizes the membrane by opening sodium and calcium channels. The entry of calcium ions induces release of the transmitter substance into the synaptic cleft. The transmitter molecules bind to receptors on the postsynaptic side of the cleft. The receptors are protein molecules with high specificity and affinity for the transmitter. Binding of transmitter to receptor affects the ion channels in the postsynaptic membrane. One of two things happens.
Either sodium channels open, some Na+ enters the cell, and the membrane potential is reduced (For example, it may fall from -70 to -60 mV.) This change is called an excitatory postsynaptic potential (EPSP), because a sufficient number of such potentials in a short time will add together and excite (depolarize) the neuron enough to initiate an action potential;
Or chloride channels (or potassium channels) open, some Cl- enters (or K+ leaves) the cell, and the membrane potential is increased (For example, it may rise from -70 to -80 mV.) This change is called an inhibitory postsynaptic potential (IPSP), because the hyperpolarization of the membrane makes excitation more difficult to achieve.
There are two general types of neurotransmitter receptor.
A ligand gated ion channel receptor has receptor and effector components in the same unit. Binding of neurotransmitter (the ligand) to the receptor part of the molecule on the outer side of the membrane induces a conformational change in sequences of amino acids that are clustered to form a gate at the inner (cytoplasmic) end of the channel. The conformational change opens the gate and allows the passage of ions. The selectivity for particular ions probably resides in the distribution and charge of basic and acidic amino acid side chains that form the open gate. Ions pass through, following the concentration gradient, if they are small enough and are appropriately charged. Ligand gated ion channel receptors evoke the most rapid postsynaptic changes.
With a G-protein coupled receptor, binding of the ligand to a specific sequence of amino acids at the external surface of the cell membrane activates a long sequence known as a G-protein at the cytoplasmic surface. G-proteins have affinity for the nucleotide GTP and they become GTPases when activated. A G-protein is also coupled to an enzyme that can set off a second messenger system, such as adenylate cyclase, phosphodiesterase or phospholipase. Activation of the G-protein triggers the second messenger system, greatly amplifying the consequences of the initial ligand-receptor interaction. The second messenger system acts upon ion channels, which may or may not be parts of the complex receptor molecule. Some G-protein coupled receptors are also channels, and they resemble ligand gated ion channel receptors in that they evoke rapid (< 10 milliseconds) postsynaptic responses. When the second messenger has to diffuse through the cytosol to some site elsewhere, the postsynaptic response is slower, and can modulate the responses of a neuron to brief but unimportant cannonades of excitatory or inhibitory input.
The postsynaptic changes in membrane potential spread laterally over the surface of the cell by electrical conduction through the cytoplasm. The dendrites and soma of a neuron ordinarily receive many synapses of both excitatory and inhibitory types, so that EPSPs and IPSPs are constantly developing at many points on the surface of the cell. The two types of postsynaptic potential have opposing effects on the net membrane potential. If the potential at the axonal hillock is reduced to a threshold value, an all-or-none action potential is initiated and travels along the axon.
The efficacy of an individual synapse is determined by its distance from the site at which impulses are initiated. Thus, a small number of IPSPs produced near the axonal hillock can counteract the effect of a large number of EPSPs in the ends of the dendrites. Hyperpolarization in the dendrites and soma of a neuron is called postsynaptic inhibition. There are axo-axonal synapses that can arrest the propagation of impulses in the terminal parts of axons, preventing the depolarization of the presynaptic terminals. Such synapses produce presynaptic inhibition.
Action potentials occur only in axons, so they give rise to EPSPs or IPSPs only at synapses where the presynaptic elements are axons. When the presynaptic neurite is not an axon, as in a dendro-dendritic synapse, the release of the transmitter is triggered by a smaller, slower depolarization of the membrane. The released transmitter evokes a postsynaptic potential, just as it would at an axo-dendritic synapse.
There are some neurons that never conduct action potentials. At the synapses of such cells, transmitters are released in response to smaller, slower reductions of membrane potential than those that occur in axons. For example, the photoreceptors of the vertebrate retina have sodium channels that leak when it is dark. The resultant lowering of the membrane potential causes continuous release of the neurotransmitter, which excites the interneurons of the retina. Light makes the sodium channels of the photoreceptors close, causing hyperpolarization and cessation of release of the transmitter. The interneurons of the retina also exhibit only slow changes of membrane potential. So do some other small neurons such as the granule cells of the olfactory bulb, whose only neurites are dendrites.
Neurotransmitters and neuromodulators
It was once thought that a neuron used only one transmitter at all its synaptic boutons. It is now known that most neurons contain at least two or three substances potentially capable of being transmitters, and in many cases the appropriate receptor molecules are also known to be present at postsynaptic sites. This knowledge has been gained mainly by immunohistochemistry, a family of staining techniques based on the use of antibodies that bind specifically to the receptors, transmitters, or enzymes of transmitter metabolism. Often drugs are available that compete specifically with the natural transmitter for binding to the receptors. Such drugs may mimic the transmitter's actions (agonists) or inhibit synaptic transmission by blocking the receptors (antagonists or blockers).
The typical action of a neurotransmitter on the postsynaptic membrane is the production of either an EPSP or an IPSP. The same transmitter will often produce both effects, though at different sites. This is because the postsynaptic response is determined not by the identity of the neurotransmitter, but by a property of the receptors to which it binds.
Two examples will illustrate this point.
1. Acetylcholine, a transmitter at many peripheral (and central) synapses, causes contraction of skeletal striated muscle cells. This action is mimicked by low concentrations of nicotine, blocked by the curare alkaloids, but unaffected by atropine. Acetylcholine also causes contraction of intestinal smooth muscle, but here the action is mimicked by muscarine, antagonized by atropine and unaffected by curare. The nicotinic receptor is ligand-gated, whereas the muscarinic receptor is G-protein coupled.
2. Noradrenaline is the transmitter used by most of the visceral (sympathetic) postganglionic neurons that supply blood vessels in skeletal muscle and skin. Because of different receptors, noradrenaline causes vasoconstriction in the skin and vasodilation in muscle.
The coexistence of two or more neurotransmitters in the same neuron allows the effect of a rapidly acting transmitter to be influenced by the actions of more slowly acting substances, called neuromodulators or co-transmitters. These may not themselves induce an EPSP or an IPSP, but rather change the properties of the postsynaptic membrane so that the response is not the same as that evoked by the fast-acting transmitter alone. Peptides and some of the amines secreted by neurons are thought to be neuromodulatory agents, though some may also be rapidly acting transmitters at other synapses. As previously pointed out, rapid changes in postsynaptic membrane potential result from activation of ligand-gated channels. Slower changes and metabolic adjustments are mediated by G-protein-couple receptors.
Table 3 is an annotated list of substances thought to be neurotransmitters or neuromodulators in vertebrates. The list is far from complete. Many of these substances are also present and active in the nervous systems of invertebrate animals.
Axonal transport
A simple calculation shows that even a small neuron has most of its cytoplasm in the neurites. A long axon accounts for over 99% of the volume of the cell. Almost all the neuron's DNA and RNA are in the soma, so it is necessary for synthesized proteins and other substances to be transported distally in the dendrites and axon. Most of the knowledge of such transport has come from the study of the movements of substances in the cytoplasm of axons, hence the term axonal (or axoplasmic) transport.
Velocities and directions of transport
Although axons are very narrow tubes, different substances move within them at different speeds, and even in different directions, at the same time. Transport away from the cell-body is anterograde; that towards the soma is retrograde. The largest amounts of material moved in an axon constitute the slow component of anterograde transport, moving at about 1 millimeter per day. The substances transported in the slow component are mostly structural proteins: tubulin (the subunits of microtubules), actin (for microfilaments), and the subunits of the neurofilament proteins. The fast component of anterograde axoplasmic transport moves at 400 mm per day in mammals and birds, or at 200 mm per day in cold blooded vertebrates. Microtubules mediate fast transport, which is prevented by the microtubule-disrupting drugs such as colchicine and vinblastine. Substances moved in the fast component are contained in particles such as mitochondria or small vesicles of the smooth endoplasmic reticulum. They include enzymes of neurotransmitter metabolism and peptides that are transmitters or neuromodulators.
Retrograde axonal transport occurs at about half the velocity of the fast anterograde component. Of the substances sent to the cell body, the most interesting are those that originate outside the neuron. Studies with radioactive and histochemically detectable tracers indicate that the presynaptic parts of an axon imbibe substances from the surrounding extracellular space. Such materials are then retrogradely transported to the soma, where they are sequestered in lysosomes and eventually degraded. Axonal uptake and retrograde transport provide a mechanism whereby the soma can receive information about the extracellular environment of remote parts of the cell by direct sampling.
Neuroanatomical tracing methods based on axoplasmic transport
The axons and dendrites in the central nervous system are usually so closely interwoven that it is impossible to determine their exact sites of origin and termination by direct observation. Methods that label specific populations of neurons in experimental animals contribute greatly, therefore, to the acquisition of neuroanatomical knowledge. Such investigations are conducted in one of the following ways:
1. A radioactively labelled amino acid is injected into the region of the somata of a group of neurons. It is incorporated into proteins, some of which are transported anterogradely to the presynaptic axonal terminals. The appropriate parts of the brain or spinal cord are removed, after allowing time for rapid transport (often 24-48 hours), and prepared for autoradiography. The silver grains in the autoradiographs show the site of the injection, the axonal terminals, and often the trajectory of the intervening axons.
2. A histochemically demonstrable enzyme, horseradish peroxidase, is injected into the region to be studied. The enzyme may, with advantage, be chemically coupled to a lectin (a protein of plant origin, with affinity for cell surfaces). After 1 to 3 days, the distribution of the enzyme is examined histochemically. The sections reveal the cell bodies of the neurons whose axons end in the site of injection, and also the axonal terminals of the neurons whose somata and dendrites were in the injected region. This method makes use of both anterograde and retrograde transport.
3. Different fluorescent dyes are injected at two different sites. After allowing 2 to 4 days for uptake by presynaptic terminals and retrograde axonal transport, sections of the tissue are examined by fluorescence microscopy. The somata sending axons to the two injected sites fluoresce in different colors. A neuron is labeled in both colors if its axon sends branches to both the injected sites.
These methods can be used in all types of animals and are applicable to any part of the nervous system. They are the most accurate tracing methods available but cannot, of course, be used in human beings.
------------- End of introductory material -------------
Section 4. Comparative neuroanatomy.
The nervous systems of two radially symmetrical animals are shown in Figure 10 - Radially symmetrical animals. In the simpler animal, Hydra, the neurons have two to five neurites each and are uniformly distributed throughout the body wall, beneath its ectoderm (outermost layer). Physical contact at any point causes the cells of the wall to contract and bend toward the stimulus. A food particle in the midst of the tentacles is thus pushed into the mouth, and simple locomotion of the animal is also possible. The contraction of cells remote from the site of stimulation is possible because communication among the neurons of the nerve net is much faster than among ordinary cells. The intensity of the stimulus determines the number of neurons that become active and thereby dictates the size of the movement. There are no preferred routes of communication within the nervous system of Hydra. Cnidaria (coelenterates) more advanced than Hydra, such as sea anemones and jellyfish, have more complicated nerve nets, with different populations of neurons coordinating different movements, and sometimes with regions that can be designated as ganglia because they are densely populated with neurons. The most advanced radially symmetrical animals are adult echinoderms, such as the starfish Asterias (Fig. 10). In these, as in the Cnidaria, there is no tendency towards a single centralized system. The neurons are, however, concentrated into rings and radiating cords.
The simplest animals with bilateral symmetry are the platyhelminths or flatworms, such as the common planarian, Dugesia (Fig. 11 - Flatworm nervous system). Flatworms resemble the Cnidaria in having a single orifice that serves as mouth and anus. In these animals there is a diffuse nerve net beneath the ectoderm, but neurons are also present in longitudinally running nerve cords. Dugesia has two such cords, connected with the superficial nerve net and, through commissures, with one another. At the anterior end of a planarian the nerve cords are enlarged and fused, forming a cerebral ganglion. This also receives input from nearby chemosensitive (taste) cells, from the light sensitive eye-spots, and from a gravity-detecting structure, the statocyst. The cerebral ganglion is the simplest kind of brain. In it, the neuronal cell bodies occupy a peripheral rind, and neurites contact one another in the central core, in a tissue known as neuropil. This rind-and-core structure is seen in the ganglia of all the higher invertebrates too, but not in the nervous systems of vertebrate animals.
The nervous systems of various worms and worm-like animals are shown in Figure 12 - Lowly and less lowly worms. These animals all have the mouth and anus widely separated and have well-developed cerebral ganglia around the pharynx and esophagus in what can be unequivocally called a head. Many zoologists think some sort of worm was the ancestor of all vertebrate animals. Candidates include echinoderms (whose larvae are bilaterally symmetrical), the nemerteans or the cephalochordates, or extinct forms resembling these. Less convincing putative ancestors that have been suggested at one time or another include annelids and scorpion-like arthropods.
The invertebrates that exhibit the most elaborate behavior are some of the arthropods and molluscs (Fig. 13 - Insect, scorpion and snail). Arthropod nervous systems conform to a general plan, with a brain and one or two ventral nerve cords with segmental ganglia. The nervous systems of molluscs vary greatly in size and complexity, as is to be expected in animals as diverse as clams, snails, and squids. Most behavioral patterns of invertebrates are "hard-wired" into the nervous system. Learning and adaptation are severely restricted by the inadequacy of connections between ganglia in different parts of the body.
The phylum Chordata consists of animals in which, at least in some stage of development, there is a tubular cord of nervous tissue dorsal to a skeletal element, the notochord. The phylum includes the worm-like amphioxus (see Fig. 12) and all vertebrate animals.
Vertebrates.
All vertebrates have a central nervous system consisting of the brain and spinal cord, protected by the axial skeleton, and a peripheral nervous system consisting of nerves and ganglia (see Fig. 1 - Generalized CNS and PNS).
Spinal and cranial nerves.
The spinal nerves are segmentally organized. The most primitive vertebrates, the cyclostomes (lampreys and hagfish), have alternating dorsal and ventral spinal nerves. The ventral nerves consist entirely of the axons of neurons in the spinal cord that supply striated skeletal muscle. The dorsal nerves, each with a spinal ganglion, contain the axons of neurons in the cord that control viscera, together with all sensory axons. A spinal ganglion contains the cell-bodies of sensory neurons, each with a distal neurite distributed in a peripheral nerve and a central neurite that enters the spinal cord.
The dorsal and ventral spinal nerves of cyclostomes correspond in all higher animals to the dorsal and ventral roots, which unite to form the mixed spinal nerves. In fishes and amphibians, efferent axons concerned with visceral function leave the spinal cord in the ventral and dorsal roots. In reptiles, birds, and mammals, however, there is complete segregation of afferent and efferent nerve fibers, with the dorsal roots containing only sensory fibers.
The cranial nerves, which are connected with the brain, have more diverse functions than the spinal nerves. The names, numbers, and principal functions of the cranial nerves are summarized in Table 4 - Cranial nerves - comparative summary.
Special sense organs.
Small groups of ectodermally derived sensory cells called neuromasts are widespread in the skin of cyclostomes and fishes. In the latter, and also in the aquatic larvae of amphibians, they occupy a system of subcutaneous canals, the lateral line system. The canals communicate through pores in the overlying skin with the surrounding water. Lateral line receptors, which are innervated by cranial nerves (V, VII, IX, X), detect low pressure changes (not sounds, which are pressure waves of higher frequency) in the water. In some fishes they also detect electric signals emitted by other fishes. Some of the neuromasts of lampreys are sensitive to light. The sensitivity of neuromasts to radiation and to mechanical stimuli indicates that these sense organs may, in ancestral vertebrates, have evolved into parts of the more advanced special sense organs of higher animals: the inner ear, the taste buds, and the infrared (radiant heat) detecting pits on the heads of some snakes. The olfactory epithelium is thought to have evolved independently of the neuromast system, because it is already well developed in cyclostomes, the most primitive living vertebrates.
Central nervous system.
The spinal cord has a hollow core of gray matter surrounded by longitudinally coursing axons. The latter include myelinated fibers in all vertebrates more advanced than cyclostomes. The central canal of the spinal cord expands rostrally into the ventricles of the brain.
The shape of the brain (Fig. 14 - Brain in 6 vertebrate groups) is due partly to the ventricular system and partly to the variable thickness of the nervous tissue forming its walls. As in man, all vertebrates have a hindbrain, midbrain, diencephalon, and telencephalon (forebrain). In fishes and amphibians, the most conspicuous thickenings are in the left and right sides of the dorsal wall of the midbrain. These constitute the optic tectum, which corresponds to the superior colliculus of the human brain. In higher vertebrates, the tectum (Latin for "roof") is relatively smaller. The cerebellum is an outgrowth of the dorsal and lateral surfaces of the hindbrain. It is present in all vertebrate groups but is largest in mammals. The optic nerve and the retina of the eye are outgrowths of the diencephalon. This part of the brain also has two glandular outgrowths: The epiphysis (pineal body or gland) dorsally, and part of the hypophysis (the neural part of the pituitary gland) ventrally.
The size and complexity of the telencephalon increase with phylogenetic advancement. The probable homologies of its major parts, the septum, the corpus striatum, and the pallium, are shown in Figure 15 - Forebrain in 8 animals. In mammals the telencephalon is larger than in other vertebrates and its most conspicuous part is the pallium, which is also called the cerebral cortex. The gray matter of the cerebral cortex is on the external surface of the cerebral hemisphere.
Variations in the structure of the mammalian brain are associated with differences in posture, behavior and intelligence. A quadrupedal animal has its eyes looking along the ground, so its brain and spinal cord form a more less straight tube. In the bipedal condition, which is most developed in man, the eyes are lined up in a plane at right angles to the axis of the spinal column; this necessitates a sharp bend in the axis of the central nervous system, formed largely from the mesencephalic and telencephalic flexures of the embryo.
Another source of variation among groups of animals is the importance of the chemical senses, especially smell (olfaction), which is used in the activities of feeding, defense and reproduction. Many mammals rely heavily on olfaction for their survival, whereas others are more dependent on their eyes and ears than on their noses. Corresponding differences in the brain reflect the relative importances of the special senses. Animals in which olfaction predominates are macrosmatic, whereas those that rely less on their noses are microsmatic. The insectivores (shrews, hedgehogs, moles etc.) and rodents (rats, mice etc.) are macrosmatic mammals. Primates (monkeys, apes, ourselves) and cetaceans (whales, porpoises, dolphins) are microsmatic. Ungulates (horses, cattle, pigs etc.) and carnivores (ferrets, cats, dogs etc.) are intermediate. The microsmatic animals are often endowed with high intelligence.
The more intelligent an animal is, the larger is its telencephalon. The cerebral cortex forms the smooth (lissencephalic) surface of the cerebral hemisphere in such animals as the rat and rabbit. Increased numbers of cortical neurons are accommodated by increasing the surface area of the hemisphere. This is achieved by folding the cortex into convexities (convolutions or gyri), separated by grooves (sulci). A brain with gyri and sulci is said to be gyrencephalic.
Why do tracts cross the midline?
A commissure contains axons that symmetrically connect regions of the two sides. Commissural neurons exist in platyhelminths and all higher animals. A decussation is a site where axons from one part of the central nervous system cross the midline on their way to a different region on the other side. Medical students often ask why so many human ascending and descending pathways include decussations. Why are the affairs of one side of the body conducted by neurons in the opposite side of the brain? Much of the mammalian cerebral hemisphere (thalamus, corpus striatum, cerebral cortex) is especially notable for its connections with the skin, muscles and visual fields of the contralateral side.
Some caudal parts of the brain, notably the vestibular nuclei and the cerebellum, have predominantly ipsilateral connections with sensory receptors and muscles. The olfactory and gustatory pathways also do not decussate. The other major sensory systems (vision, hearing, touch, conscious proprioception) include great numbers of fibers that cross the midline on their way to the forebrain. Descending pathways (from the forebrain to motor neurons in the brain stem and spinal cord) are also largely crossed. To compensate for their ipsilateral vestibular and proprioceptive input, the left and right halves of the cerebellum communicate with the contralateral thalamus and cerebral cortex by way of pathways that include long, decussating axons.
Comparative neuroanatomists cite decussations as an example of the continued exploitation of a structural feature that helped our lowly ancestors escape from predators more efficiently than their even more lowly competitors. Natural selection would not allow the loss of a decussating pathway if this were an advantage in a world full of other edible animals with non-decussating neural connections. In order to have left and right sides an animal must have different dorsal and ventral surfaces. The struggle for survival is supposed to have been among animals that lived where "dorsal" and "ventral" were significantly related to he surroundings (on the ocean floor,in shallow water, or on land). Even the most primitive nervous systems include motor and sensory neurons. A potentially fatal stimulus should evoke a movement of withdrawal, so that the attacked individual may survive and reproduce itself. The animal is more likely to escape by moving away from the assaulted side, especially if the predator is not smart enough to predict such a response. The fastest neuronal circuit for stimulating withdrawal to the other side of the midline is a monosynaptic reflex: a sensory neuron has an axon that crosses the midline and contacts motor neurons that make nearby muscle fibers contract. Such an arrangement makes a worm-like creature bend away from the attacked side. It may be significant that even in man the midline is crossed at a more caudal (presumably more primitive) level in the pathway for pain than in the pathway for discriminative sensations. These evolutionary speculations suggest that it is generally advantageous to have neurons capable of sending their axons across the midline.
Decussating pathways in vertebrates allow for the formation of congruent representation in the brain of images in the visual fields of the two eyes. The camera-type eye of vertebrate animals projects an inverted image onto its retina, so that events in the left half of the visual field of the left eye trigger neural signals that arise in the right half of its retina. If these signals were sent to the right side of the brain, the inverted projection would be a mirror image of the equivalent projection from the right half of the visual field (Fig. 16 -Uncrossed visual pathway).
A decussating projection from the retina to the brain assures that the central topographic representations of the visual fields are correctly adjacent. In most vertebrates, the eyes see separate visual fields, and all the fibers of the optic nerve cross the midline (Fig. 17 - Completely decussating visual pathway). Some mammals (including man) have forward-facing eyes with overlapping visual fields. In this case the decussation of only the fibers from the medial half of the retina provides a correctly aligned topographical projection to the brain (Fig. 18 - Partial visual decussation).
A visually guided movement is most likely to be needed on side from which the visual signal originates. Projection of the left and right visual fields to the contralateral tectum or cerebral hemisphere provides for rapid communication between the visual pathway and the motor neurons that work the muscles of the opposite side of the body. The visuomotor connections are ipsilateral in the brain but require a compensating decussation in the tracts that descend to the motor neurons (Fig. 17).
Section 5. Development of the nervous system
situated, as the neural folds. Thus, a neural groove is formed along the length of the dorsal surface of the embryo. The groove deepens, and the neural folds come into contact with one another and fuse, so that a neural tube is formed. The fusion occurs first in the middle part of the neural groove, destined to become the lower cervical segments of the spinal cord, on about the 22nd day after fertilization. Fusion of the neural groove proceeds rostrally and caudally (Fig. 19 - Human embryo, 21 days). The neural tube sinks into the mesoderm, and the continuity of the ordinary ectoderm, which will become the epidermis, is restored. The holes at the ends of the neural tube are the rostral ("anterior") and caudal ("posterior") neuropores. In man, the rostral neuropore is closed off by continued growth of the neuroectoderm on the 24th day after fertilization. The caudal neuropore closes on the 27th day.
The neural tube will become the central nervous system. The site of closure of the rostral neuropore is represented in the adult brain by the lamina terminalis, in the rostral wall of the third ventricle. The site of closure of the caudal neuropore corresponds to the upper lumbar level of the spinal cord. In man, the segments of the spinal cord caudal to L2 are not derived from the neural tube but from the caudal cell mass. This is a population of neuroectodermal cells that develops between the caudal end of the neural tube and the embryonic tail. Holes that develop in the caudal cell mass fuse with one another and eventually (about Day 48) with the caudal end of the neural tube. The growth of the rostral end of the neural tube to form the brain is described later.
Neural crest
Some of the cells of the neural folds are left behind near the dorsal surface of the embryo when the neural groove closes to form the tube. These cells constitute the neural crest. They proliferate and migrate extensively through the mesoderm, giving rise to the neurons and glial cells of the peripheral nervous system and to several other tissues. Non-neural derivatives of the neural crest include melanocytes of the skin,
various endocrine cells, and some of the bones, muscles and connective tissue of the head, including. The morphogenetic movements that produce the neural tube and neural crest are summarized in Figure 20 - Neural tube and crest diagram.
Placodes
A few parts of the peripheral nervous system are derived from placodes, which are localized thickenings of the ectoderm in the head region. Thus the olfactory epithelium develops from an olfactory placode, and placodes also give rise to some of the neurons in the sensory ganglia of cranial nerves V, VII, IX, and X. Other placodes form the lens of the eye and the sensory epithelium and associated neurons of the inner ear.
Formation of the brain and spinal cord
Histogenesis
The wall of the neural tube has three layers. The ventricular layer is next to the lumen, and all mitoses occur in this layer. External to the ventricular layer is the mantle layer, consisting of cells descended from those of the ventricular layer. The marginal layer is the outermost; it consists largely of neurites. The cells in the mantle layer that become neurons are called neuroblasts; those that differentiate into neuroglial cells are glioblasts. In the spinal cord and brain stem, the marginal layer eventually comes to consist largely of masses of myelinated axons and their supporting glial cells, a tissue named white matter, from the color of the myelin. The mantle layer develops into masses of neuronal somata, dendrites, and synaptic connections. The color of such tissue in preserved anatomical specimens has given origin to the term gray matter, even though in the living state it is pink, owing to its rich blood supply. In the cerebrum, the cortex contains neurons derived from the mantle and marginal layers. The ependyma lining the ventricles and central canal is the remnant of the ventricular layer of the neural tube.
Spinal cord, brain stem, and cerebellum
The architecture of the neural tube remains recognizable in the adult spinal cord. The alar plate, consisting of the ventricular and mantle layers of the dorsal part of the tube, becomes the dorsal horn of the spinal gray matter. The basal plate in the ventral part of the tube becomes the ventral horn (Fig. 21 - Alar and basal plates). Neurons derived from the alar plate receive synapses from the primary sensory neurons in the spinal ganglia, whereas the neurons from the basal plate include the motor neurons that innervate striated skeletal muscle. The alar and basal plates give rise to equivalent sets of neurons in the brain stem, even though this part of the neuraxis develops a shape quite different from that of the spinal cord (Fig. 21). The roof-plate of the medulla and pons is wide and thin. Consequently the alar plate lies laterally, and the basal plate lies medially, in the floor of the fourth ventricle. The sulcus limitans, separating the alar from the basal plate, persists in the floor of the adult human fourth ventricle. Groups of motor and autonomic neurons lie medial to the sulcus limitans; groups of neurons lateral to the sulcus receive the incoming axons from the sensory ganglia of cranial nerves. The midbrain retains a tubular form, with most of the white matter in its ventral and lateral parts, and gray matter dorsal and ventral to the aqueduct.
The cerebellum arises as an outgrowth from the dorsolateral aspects of the brain stem. It has deep gray matter, associated with the rostal part of the roof of the fourth ventricle, and a cortex of gray matter on the external surface.
Diencephalon and telencephalon
The rostral part of the neural tube grows much more than the parts that give rise to the brain stem and spinal cord, and flexures develop so that the brain comes to fill the cranium. The result is the formation of the cerebral hemispheres (Fig. 22 - Development of cerebral hemispheres). The arrangement of central gray matter and external white matter is conserved in the cerebral hemispheres, but not in an obvious way. The gray masses of the thalamus and hypothalamus flank the third ventricle, and external to them is the internal capsule, a band of white matter of great functional importance. The lateral ventricles have large gray masses, the corpora striata in their walls, with the centraL white matter of the hemispheres further out. The development of the cerebral hemispheres is complicated, however, by the formation of the cerebral cortex. This is formed from neurons that migrate from the ventricular layer to the outside surface of the developing brain, As a result, most of the outside surface of the adult hemispheres consists of the gray matter of the cerebral cortex. The corpus callosum, a large mass of axons interconnecting the cortices of the two hemispheres, forms the roofs of both lateral ventricles.
Formation of the peripheral nervous system
Some neural crest cells form aggregates close to the neural tube and become the spinal ganglia. Other groups of neural crest cells migrate ventrally to form the ganglia of the autonomic nervous system and the sensory ganglia of cranial nerves. The enteric nervous system is derived largely from the neural crest cells that also form the vagus nerve. The cells giving rise to ganglia differentiate into both neurons and neuroglial cells. Other cells from the neural crest travel further to form the neuroglia (Schwann cells or neurolemmocytes) of all the peripheral nerves. The contribution of placodes to the peripheral nervous system has already been mentioned.
The most notable activity of neural crest cells is their migration, which has been intensively studied in recent years by making chick-quail chimeras. This is done by replacing a small piece of a chick embryo with a corresponding bit of tissue from a quail embryo. Development is then allowed to continue. The quail cells are incorporated into the growing chick, but can be recognized by virtue of a conspicuous lump of DNA in the interphase nucleus. This is absent from chick cells. It is reasonable to extrapolate to mammals from the results of experiments with chick-quail chimeras, because the conclusions correspond closely to those derived from "static" observations made on graded series of embryos of different ages. The anatomical organization of the peripheral nervous system is, in any case, similar in all vertebrates.
Special sense organs
The organs of smell, sight, taste, hearing, and equilibration are all derived, at least in part, from ectodermal placodes rather than from the neuroectoderm.
The olfactory placode, which is part of the larger nasal placode, forms the olfactory epithelium. This consists of neurons and supporting cells that are always in contact with the external environment. The axons of the olfactory neurons grow towards and enter the most rostral part of the embryonic forebrain. Even in adult mammals, the neurons of the olfactory epithelium have limited life spans and are replaced from a population of undifferentiated precursor cells.
The development of the eye (Fig. 23 - Development of eye) is
a process of great complexity. Initially an outgrowth from the
diencephalon forms the optic stalk, dilated at its end to form an optic
vesicle. Invagination of the vesicle forms a two-layered optic cup,
which will become the retina. When the optic vesicle touches the
ectoderm on the surface, a placode is formed and it sinks in towards
the developing neural components of the eye. It will become the lens.
Mesodermal cells, attracted into the fissure between the developing
optic cup and the lens placode, eventually form the choroid, iris,
sclera, vitreous body, and blood vessels. The eye is largely formed by
the end of the 7th week after conception.
Click here for Fig. 23A, a more
detailed picture of early development of the eye.
Taste cells are derived from the ordinary ectoderm of the mouth. They form small clusters within the epithelium and acquire their chemical sensitivity when they are contacted by the growing axons of neurons in the sensory ganglia of the VIIth, IXth or Xth cranial nerves. Small groups of the innervated cells constitute taste buds. Most of them are on the tongue. Curiously, taste buds are more widely distributed and much more numerous in late fetal life than after birth.
The otic placode appears 3 weeks after conception; it sinks into the mesoderm and forms the otic vesicle, which will become the sensory epithelia of both the cochlea (for hearing) and the vestibular labyrinth (for equilibration), as well as the sensory ganglia of the cranial nerve (VIII) that serves the sensory cells. The neuroglia of the vestibulocochlear nerve originates from the cranial neural crest. The middle ear is an endodermal derivative, being an outgrowth of the pharynx. The ossicles are mesodermal. The external ear is a late development, the external meatus opens in the 6th month of intrauterine life, and the growth of the pinna continues for years after birth.
Developmental abnormalities of the central nervous system
The most serious abnormalities to arise during development of the nervous system are due to failure of closure of the neural groove. Nervous tissue is exposed on the dorsal surface of the body, because there is also failure of development of the overlying meninges, vertebral arches, and skin. If the neural groove does not close at its cranial end, the whole forebrain and the overlying skull and scalp will be missing, although the face and eyes will be formed. This condition is called anencephaly (meaning "no brain"), and the afflicted individual is either stillborn or survives for only a few hours. The corresponding condition at the caudal end of the central nervous system is myeloschisis ("split spinal cord"), with extensive exposure of non-functional nervous tissue in the lumbosacral region. Myeloschisis and anencephaly often occur together.
Myeloschisis is the most severe form of spina bifida, a condition in which there are missing vertebral arches, nearly always in the lumbar region. In the less severe varieties, the spinal cord and its immediate coverings (the leptomeninges) are formed and there is a covering of skin, but the associated mesodermal structures are defective. Thus there may be a meningomyelocele, which is a swelling containing nerve roots, spinal cord and leptomeninges protruding through the hole formed by the missing vertebral arches and dura. (The dura is the strong collagenous membrane that lines the cranium and spinal canal.) If the spinal cord amd nerve roots remain near the vertebral bodies, there is a cystic swelling composed of leptomeningeal tissue and cerebrospinal fluid. This is a meningocele. The least severe of these conditions is spina bifida occulta, in which the dura is intact but one or more vertebral arches are missing. Spina bifida occulta may cause no symptoms, or it may be blamed for otherwise undiagnosed pain in the back. Meningomyelocele and meningocele can be corrected surgically, but often the child remains paraplegic (lower limbs paralyzed). Meningomyelocele is often associated with internal hydrocephalus, a condition in which an excessive volume of cerebrospinal fluid accumulates because of an obstruction to its flow through the ventricular system of the brain.
Section 6. Some illustrations of animal and human neuroanatomy
| Figures 24-37 all have labels & legends. | ||
| Fig. 24 | Brain stem, cerebellum in rat, sheep, cat. | Click here for picture |
| Fig. 25 | Rat brain, dorsal view. | Click here for picture |
| Fig. 26 | Rat brain, ventral view. | Click here for picture |
| Fig. 27 | Rat brain stem etc., dorsal exposure. | Click here for picture |
| Fig. 28 | Medial dissection, rat hippocampus. | Click here for picture |
| Fig. 29 | Cat: lateral hemisphere. | Click here for picture |
| Fig. 30 | Cat: medial hemisphere. | Click here for picture |
| Fig. 31 | Cat cerebrum, posterior view. | Click here for picture |
| Fig. 32 | Dog brain, dorsal view. | Click here for picture |
| Fig. 33 | Dog cerebrum, lateral view. | Click here for picture |
| Fig. 34 | Dog cerebrum, medial view. | Click here for picture |
| Fig. 35 | Human brain stem, lateral & ventral views. | Click here for picture |
| Fig. 36 | Human brain stem, 4th ventricle, dorsal view. | Click here for picture |
| Fig. 37 | Human diencephalon; medial view & 2 coronal sections. | Click here for picture |
| Figures 38-50 are labelled transverse sections of the rat's brain, at approximately 1.5 mm intervals. | ||
| Fig. 38 | Section just caudal to olfactory bulbs. | Click here for picture |
| Fig. 39 | Rostral cerebrum with anterior part of septum. | Click here for picture |
| Fig. 40 | Rostral cerebrum; middle of septal region | Click here for picture |
| Fig. 41 | Forebrain, posterior septal region. | Click here for picture |
| Fig. 42 | Diencephalon; also hippocampal commissure. | Click here for picture |
| Fig. 43 | Diencephalon; amygdala, hippocampus. | Click here for picture |
| Fig. 44 | Pretectal level. | Click here for picture |
| Fig. 45 | Midbrain, Superior colliculi; also pons. | Click here for picture |
| Fig. 46 | Pons; inferior colliculi. | Click here for picture |
| Fig. 47 | Medulla/pons; VI & VII nuclei. | Click here for picture |
| Fig. 48 | Rostral medulla; all 3 cerebellar nuclei. | Click here for picture |
| Fig. 49 | Mid-medulla. | Click here for picture |
| Fig. 50 | Caudal medulla; corticospinal decussation. | Click here for picture |
Interactions of larger parts of the nervous system are understandable only in terms of the activities of large numbers of individual cells. The definitions presented in Table 5 will remind the reader of (a) the terminology of Neuroscience relating to communication within and among cells, and (b) the words used to specify parts of the nervous system on a larger scale. For graduate students in the basic biomedical sciences, Table 5 may be useful as a glossary of human neuroanatomical terms that are often applied (or misapplied) to the nervous systems of mammals generally.
The formation of a spinal nerve is illustrated in Fig. 51 - Spinal nerve. This diagram also shows structural elements that will be referred to later in this Section. Spinal nerves have numbers derived from the vertebrae. The highest spinal nerve penetrates the atlanto-occipital membrane, above the arch of the atlas, which is the first cervical vertebra or C1. The second cervical nerve passes between the atlas (vertebra C1) and the axis (C2). There are 7 cervical vertebrae. The lowest cervical nerve is therefore C8. Cervical nerves 1 to 7 go through foramina above the numbered vertebrae. The roots of nerve C8 pass below the arch of vertebra C7 and above that of T1. All the thoracic (T1 - T12), lumbar (L1 - L5) and sacral (S1 - S5) nerves go through foramina below the equivalently numbered vertebrae. To complete the story, a single coccygeal nerve overlaps with S5 in supplying the perianal skin.
The most obvious consequence of the segmental organization of the spinal nerves is seen in the Dermatomes, which are bands of skin that run horizontally on the trunk and lengthwise on the limbs (Fig. 52a - Dermatomes). Each dermatome is centered on the distribution of axons from a single dorsal root ganglion, but each ganglion also supplies skin in the dermatomes above and below its own level. Consequently, it is necessary to transect three adjacent dorsal roots or spinal nerves in order to completely denervate the skin of one dermatome. Transection of a single spinal nerve, or destruction of its ganglion, diminishes but does not abolish sensation in the affected segment of skin. The cutaneous lesions of herpes zoster, a common virus that infects certain pain-responsive neurons in individual sensory ganglia, often neatly map the distributions of dermatomes and also illustrate the extension of innervation into the adjacent segments of skin. The nerve supply to the skin of the limbs is delivered by cutaneous nerves that are formed in limb plexuses (brachial and lumbosacral) by complex interchanging and mixing of fibers from different spinal roots. The areas supplied by cutaneous nerves bear little resemblance to the dermatomes. They are sharply demarcated, with little or no territorial overlapping (Fig. 52b - Dermatomes). The widely overlapping dermatomes cut across adjacent areas of skin supplied by cutaneous nerves. A cutaneous nerve lesion, such as an injury or a mononeuropathy, results in a well defined area of defective sensation, and anatomical knowledge can be used to identify the affected nerve.
Most of the skin of the head is supplied by the three divisions of cranial nerve V. The areas are sharply demarcated, and therefore do not correspond to dermatomes. Cranial nerves VII, IX and X supply small, overlapping areas of skin of the external ear, and the dermatome of the second cervical nerve includes parts of the head, ear, face and neck. (The first cervical nerve lacks a dorsal root in most people.)
Muscles receive motor and sensory innervation. Most of the muscles of the limbs are supplied nerves formed in the limb plexuses from two or more roots. Table 6 (Segmental landmarks) shows the segmental innervation of a few clinically important muscles. A stretch reflex (tendon jerk) requires the integrity of both the motor and the proprioceptive sensory innervation of the muscle.
Relation of spinal cord and nerve roots to the vertebral column
The vertebral column is longer than the spinal cord, which ends at the level of the upper border of vertebra L3 in the newborn and at the upper border of vertebra L2 in the adult. The lower spinal nerves must therefore course caudally before passing through their corresponding intervertebral foramina. Immediately below the caudal end of the spinal cord, the neural canal contains the roots of nerves L2-L5, S1-S5 and the coccygeal nerve.
A lesion arising from the axial skeleton, such as a herniated intervertebral disk or tumor tissue from a vertebral body or pedicle, can press on the spinal cord or spinal nerves. The consequences depend on the level of the involved disk or vertebra. In the cervical and upper thoracic spine there is little discrepancy between the spinal segments and the vertebrae. There is little free space in this part of the neural canal, so a lesion is likely to impinge on the cord as well as on a spinal nerve. The body of vertebra T10 is level with spinal cord segment T11. Below this level, the discrepancy between vertebral and spinal levels increases rapidly, because the lower lumbar and the sacral segments of the spinal cord are much shorter than the cervical and thoracic segments. All the spinal cord segments below T11 are in the range of just three vertebrae, T12, L1 and L2.
The foramina are above the levels of the intervertebral disks. Consequently, a herniated disk below C7 cannot compress its own segmental nerve; it presses on the nerve one or two segments lower. For example, an L4-5 disk herniation commonly compresses spinal nerve L5 or S1, causing pain and other sensory abnormalities in the appropriate dermatomes (see Fig. 52a).
Cranial nerves
Although the brain stem develops from segments (known as neuromeres), their peripheral distributions and central connections are most easily understood in terms of the functions of each nerve. These are set out in Table 7 - Cranial nerve components. Note that the second cranial "nerve," despite its traditional name, is not a nerve but an outgrowth of the brain, as is the retina.
Knowledge of the functional components and central connections
of the cranial nerves is essential for the understanding of many of the
normal activities of the nervous system and for the diagnosis of
clinical conditions, especially those that affect the brain stem.
Loss of function of one or more cranial nerves generally indicates the level
of a lesion in the brain stem.
Loss of function in motor or sensory pathways can indicate the position
of a lesion (medial, lateral, ventral, dorsal) at the level determined
from cranial nerve involvement.
These notes do not include accounts of the individual cranial
nerves. You will need a textbook. The cranial nerves (other than those
devoted to special senses) are described in Chapter 8 of Barr's
The Human Nervous System: An Anatomical Viewpoint
(by the author of this web site). For more scholarly expositions of the
subject books by Brodal are strongly recommended. The
Central Nervous System by Per Brodal is an
excellent textbook of human neuroanatomy that covers the cranial nerves
thoroughly. The author's father, Alf Brodal, also wrote excellent
textbooks, including The Cranial Nerves,
a small paperback that is hardly out of date 30 years after its last
edition. These older books are in libraries and (if you are lucky)
second-hand bookshops.
Cranial nerves have many branches, some of which carry sensory and
sympathetic axons through parasympathetic ganglia. This is a field
where gross anatomy and neuroanatomy overlap. Neuroanatomy textbooks
for medical students usually assume a prior knowledge of the anatomy of
the cranial nerves, as revealed by dissection. Students of neuroanatomy
without prior education in human or animal anatomy will need to consult
appropriate textbooks. Green's Anatomy of the Rat
is still the best book for rodent anatomy. The pictures in the 1935
edition are superior to those in recent reprints. There are many
textbooks of human anatomy.
Autonomic nervous system
Skeletal muscles are supplied by motor neurons whose cell bodies are in the spinal cord (anterior horn) or brain stem (motor nuclei of cranial nerves). In contrast, glands, cardiac muscle, and the smooth muscle of blood vessels and internal organs are supplied by neurons in ganglia of the autonomic system. These ganglia receive afferent preganglionic fibers, which are the thinly myelinated axons of neurons in the spinal cord or brain stem. The neurons in the ganglia have unmyelinated axons, the postganglionic fibers that innervate smooth and cardiac muscle and secretory cells. There are three divisions of the autonomic system: sympathetic, parasympathetic and enteric.
The ganglia of the sympathetic system are the chains of paravertebral ganglia that lie on the lateral aspects of the bodies of the vertebrae, and also the preaortic or collateral ganglia associated with the branches of the aorta that supply abdominal organs. There is a sympathetic chain ganglion for every spinal nerve. Postganglionic fibers enter the nerve by way of a gray ramus communicans (See Fig. 51 - Formation of spinal nerve) and are distributed to blood vessels, sweat glands and the little muscles that move hairs. Blood vessels of the skin constrict in response to their sympathetic supply, whereas those in muscles dilate. Some of the ganglia for the nerves C1 to T1 are fused; consequently there are only three cervical sympathetic ganglia. In most people there is a stellate ganglion, formed from inferior cervical (C7-C8) and first thoracic ganglia. The middle cervical ganglion is connected with nerves C5 and C6. Postganglionic fibers from the large superior cervical ganglion (C1-C4) accompany the carotid artery and its branches. Some enter the eye, where they supply the dilator pupillae muscle of the iris. Others supply smooth muscle within the upper eyelid1. All three cervical ganglia send postganglionic fibers into cardiac nerves, which run alongside the common carotid artery and aorta and supply the muscle of the heart. Increased activity of the sympathetic system increases the rate and force of contraction of the heart.
| In the absence of active sympathetic innervation of the eye, orbit and face, the pupil constricts (unopposed parasympathetic action), the upper eyelid droops partly, but can be raised voluntarily (intact oculomotor nerve innervation) and sweating does not occur on the affected side of the face. These changes (Horner's syndrome) can result from damage to the superior cervical ganglion, its pre- or postganglionic fibers, (which pass through the stellate ganglion), or to transection of descending axons in the lower brain stem or spinal cord that control the preganglionic neurons. |
Parasympathetic ganglia are found in the head, connected with certain cranial nerves, and associated with the walls of thoracic and pelvic viscera. Preganglionic fibers leave the brain stem in cranial nerves III, VII, IX and XI and terminate in cranial parasympathetic ganglia. The neurons in these ganglia supply the structures whose functions are stated in Table 7 - Cranial nerve components. The cardiac ganglia receive their preganglionic afferents from the vagus nerve; their neurons supply cardiac muscle cells, principally in the atria. The pelvic splanchnic nerves, branches of S2, S3 and S4, carry preganglionic fibers to the parasympathetic ganglia that supply the detrusor muscle of the urinary bladder and the blood vessels of erectile tissue in the genitalia.
The enteric nervous system consists of thousands of tiny, interconnected ganglia in the walls of the alimentary canal, from esophagus to anus, and of some of its associated structures such as the biliary system and pancreas. These ganglia, which supply the smooth muscle and secretory tissues of the gut, contain several types of neurons, with a wide variety of neurotransmitters. The enteric nervous system can do much of its work independently, but it is modulated by preganglionic fibers from the vagus nerve (to the stomach, small intestine and first half of the colon) and from the pelvic splanchnic nerves (distal colon and rectum). Parasympathetic activity stimulates propulsion of the contents of the gut. Of the vagal fibers that enter the abdomen, a majority end in enteric ganglia of the stomach, and the integrity of this preganglionic supply is essential for acid secretion and for opening of the pyloric sphincter.
Most of the postganglionic sympathetic fibers from the preaortic ganglia synapse with neurons in enteric ganglia, but some contact blood vessels and a few supply intestinal smooth muscle. Activity of the sympathetic system causes constriction of visceral blood vessels and retards propulsion of the contents of the alimentary canal.
Regional anatomy of the central nervous system
The cerebral cortex, which covers much of the surface of the brain, is often considered to be the seat of consciousness and thinking. It receives sensory pathways, interprets the sensations, formulates commands and sends orders through motor pathways to the muscles. Although this simplistic view of neural organization is not entirely correct it has served neurologists well for more than a century, and it is still a useful starting point for anyone starting to study functional neuroanatomy. From caudal to rostral, the major divisions of the central nervous system are the spinal cord, the hind-brain (medulla, pons and cerebellum), the midbrain and the forebrain or cerebrum, which consists of two cerebral hemispheres. (see Fig. 53 - Brain surface landmarks). The brain stem comprises the medulla, pons and midbrain. The cerebellum is joined to each part of the brain stem by paired peduncles of white matter. There are therefore 6 cerebellar peduncles. The ventral part of the midbrain, on each side, is called a cerebral peduncle, because its ventral part contains great numbers of fibers descending from the cerebral hemisphere. Fig. 53 also shows some other external landmarks, which will be mentioned later.
Spinal cord
The neural components of the spinal cord are most easily understood in a transverse section through segment T1, which is connected with the nerves of the upper limb. All the ascending and descending tract are present at this level, and so are certain cell columns that occur only in the thoracic and upper lumbar segments (Fig. 54 - Spinal cord: cell columns and tracts). The small central canal of the spinal cord, which contains cerebrospinal fluid, is a remnant of the lumen of the embryonic neural tube.
The effects of a destructive lesion in the spinal cord can be predicted from knowledge of the segmental level and the functions of the tracts shown in Fig. 54. For example, A penetrating injury that transects the left half of the spinal cord in segment T4 will cause paralysis of abdominal and lower limb muscles on the left, loss of discriminative touch and proprioceptive sensation everywhere below the level of the nipple on the left side, and loss of pain and temperature sensibility below the level of the nipple on the right side.
Brain stem
The central canal of the spinal cord continues through the caudal half of the medulla and then widens to form the diamond-shaped fourth ventricle, which is the cavity of the upper medulla and of the pons. The roof of the fourth ventricle is very thin; cranial nerve nuclei form much of the floor of this ventricle, and the inferior and superior cerebellar peduncles form its walls. Cerebrospinal fluid flows from the ventricular system of the brain into the subarachnoid space through apertures in the thin roof of the fourth ventricle. At the rostral border of the pons the fourth ventricle becomes the cerebral aqueduct, which is the narrow, tubular cavity of the midbrain. Dorsal to the aqueduct is the tectum ("roof"), a substantial slab of gray matter organized into four "little hills:" the paired inferior and superior colliculi, which form parts of the pathways for hearing and eye movements, respectively.
Some of the fiber tracts of the spinal cord extend also throughout the brain stem; others begin or end there. The brain stem also contains the nuclei of cranial nerves III-XII. The motor nuclei correspond to cells of the ventral horn of the spinal gray matter, and some sensory nuclei correspond to the dorsal horn. Other cranial nerve nuclei have no obvious equivalence to cell columns of the spinal cord. Other major groups of neurons in the brain stem include nuclei connected with the cerebellum and the several cell groups that comprise the reticular formation. Fig. 55 - Tracts etc in brain stem shows the approximate levels of cranial nerve nuclei and the positions of some tracts. The interested reader will need to consult a textbook of neuroanatomy for more detailed information.
The most ventral parts of the midbrain, pons and medulla contain large numbers of fibers from the cerebral cortex that have predominantly motor functions: the corticopontine and corticospinal tracts. There are also corticobulbar fibers, which end in or near the motor nuclei of cranial nerves in the "bulb" (medulla and pons).
Two of the cranial nerve nuclei are greatly elongated. Each is a column of neurons alongside a tract, which is composed of afferent fibers from a sensory ganglion. The spinal trigeminal nucleus extends downward from a mid-pontine level until it blends with the dorsal horn of the spinal gray matter. It receives the general sensory fibers from the head and the upper end of the gastrointestinal and respiratory tracts; most of these enter the brain stem through the trigeminal and glossopharyngeal nerves. The caudal end of the spinal trigeminal nucleus receives afferents concerned with pain and thermal sensations. The rostral end is concerned with touch, as is the pontine trigeminal nucleus. The solitary nucleus, which extends the length of the medulla, receives afferent fibers from cranial nerves VII, IX and XI. The afferents to the rostral end of the nucleus are sensory neurons that innervate taste buds. The caudal end of the solitary nucleus receives signals from sensory receptors in the heart, carotid sinus, carotid body, lungs and other internal organs; it is concerned with physiological regulation of the circulatory and respiratory systems.
A large destructive lesion is the brain stem is fatal. A small, circumscribed lesion interferes with the functions of the transected tracts and of nuclei and fibers of cranial nerves. Cerebellar symptoms are present, ipsilaterally, with lesions that erode the superior or inferior cerebellar peduncle.
Cerebellum
The cerebellum consists of a midline portion, the vermis, flanked by the two cerebellar hemispheres. It has a convoluted cortex, white matter, and central nuclei. All cerebellar afferent fibers branch to end in the nuclei and cortex. The cortex projects to the underlying nuclei, which are the source of the efferent fibers of the cerebellum. As stated earlier, three peduncles on each side connect the cerebellum with the brain stem.
The best known action of the cerebellum is to ensure the correct extent and timing of movements, though additional functions have also been postulated. Cerebellar afferents come from many parts of the central nervous system, the best understood being the spinal cord, the vestibular nuclei and certain other nuclei in the brain stem, notably the inferior olivary and pontine nuclei. The cerebellum is conveniently divided into three anatomically distinct functional parts, on the basis of its afferent connections.
The vestibulocerebellum is the smallest division, with a small midline component, the nodule, and connected parts of each hemisphere, the flocculi. Afferent fibers are from the vestibular ganglion and the vestibular nuclei, which are in the rostral part of the medulla. The cortex of the vestibulocerebellum projects to the fastigial nuclei, which are embedded in the white matter of the vermis, above the roof of the fourth ventricle. The neurons in the fastigial nucleus have axons that enter the medulla and end mainly in the vestibular nuclei. These connections make the functions of the vestibulocerebellum inseparable from those of the vestibular system. Similar clinical manifestations (vertigo, nystagmus, nausea) follow damage the vestibular apparatus, nerve, nuclei or vestibulocerebellum, or to the inferior cerebellar peduncle, which carries the connecting fibers.
The spinocerebellum consists of the vermis and adjacent cortex of much of the rostral part ("anterior lobe") of the cerebellum. It receives the spinocerebellar tracts (Fig. 54) and the related cuneocerebellar tract, which relays proprioceptive signals from the upper limb. The cortex projects to the underlying interposed nuclei, and these in their turn send axons through the superior cerebellar peduncle to the contralateral red nucleus (in the midbrain) and thalamus. The spinocerebellum is driven by proprioceptive input, so if there is a lesion in the midline of the upper part of the cerebellum the motor systems fail to respond quickly to signals coming from muscles, tendons and joints. This results in poorly controlled movement (cerebellar ataxia), especially of the muscles of the trunk and lower limbs.
The pontocerebellum is the largest division, comprising most of the hemispheres and the posterior part of the vermis. The middle cerebellar peduncle, the largest, consists entirely of fibers from the contralateral pontine nuclei (Fig. 55), and the decussating pontocerebellar fibers account for the appearance and size of the ventral part of the pons. The pontine nuclei receive their afferents from extensive areas of the cerebral cortex. The deep nucleus of the pontocerebellum is the dentate nucleus, in the center of the hemisphere, and this send axons through the superior cerebellar peduncle to the thalamus, along with the efferents from the interposed nuclei. The ventral lateral thalamic nucleus, which receives the fibers from the cerebellum, projects to the primary motor area of the cerebral cortex. Thus, the pontocerebellum is influenced by activity in most of the contralateral cerebral cortex, and modulates movements by acting upon the primary motor area. Pontocerebellar disorders can be due to lesions in the dentate nucleus or in the white matter of the cerebellar hemisphere or superior cerebellar peduncle. The clinical signs, which are ipsilateral, affect the accuracy of performance of movements and include the classical manifestations of cerebellar disorder, such as dysdiadochokinesis, past-pointing and intention tremor.
It remains to be stated that all areas of the cerebellum receive input from the inferior olivary nuclei of the medulla. Afferents to the inferior olivary complex of nuclei are from the motor areas of the cerebral cortex and from the spinal cord. Physiological studies in animals indicate that the cerebellum uses its olivary afferents when learning patterns of instructions for carrying out movements. The more specific connections, described in the preceding paragraphs, are put to use in the execution of the learned patterns.
Cerebral hemisphere
The structural organization of the cerebrum and its connection with the brain stem are best appreciated in a frontal (coronal) section of the brain that passes through the ventral part of the midbrain (Fig. 56 - Coronal section of cerebrum and brain stem).
The rostral end of the midbrain merges with the diencephalon ("between brain"), which has four subdivisions on each side, separated by the third ventricle, which is a slit-like cavity in the midline.
The subthalamus, which is closest to the midbrain, contains the subthalamic nucleus, which is involved in motor circuitry, and ascending tracts that are about to terminate in the thalamus: the medial lemniscus, spinothalamic tract and fibers from the cerebellum.
The hypothalamus is medial and rostral to the subthalamus, and has landmarks on the inferior (ventral) surface of the brain. This region controls important autonomic and endocrine functions. Neural and vascular links from the hypothalamus control the pituitary gland.
The thalamus is the largest part of the diencephalon. It forms much of the wall of the third ventricle and floor of the lateral ventricle. It's many constituent nuclei communicate with the cerebral cortex. Most thalamic nuclei also receive input from subcortical sources and some are stages in pathways for sensory, motor and cognitive activities. At the anterior (rostral) end of each thalamus, the third ventricle becomes continuous with the lateral ventricle, through the interventricular foramen of Monro.
The Epithalamus is a poorly understood region of the brain associated with the junction of the cerebral aqueduct and third ventricle. It includes the pineal gland, a much studied but still mysterious probable endocrine organ, which is dorsal to the superior colliculi.
Only one large body of white matter links brain stem and diencephalon with the cerebral cortex. This is the internal capsule (Fig. 56). It consists largely of ascending thalamocortical fibers and fibers descending from the cortex to the brain stem and spinal cord. The posterior limb of the internal capsule (Fig. 57 - Horizontal section of hemisphere) includes corticospinal, corticobulbar and corticoreticular fibers with important motor functions.
The telencephalon ("end-brain") is associated with the lateral ventricle. Its central gray matter, the corpus striatum comprises the large caudate and lentiform nuclei, which will be discussed in connection with the control of movement.
The external surface of the telencephalon is formed by the cerebral cortex. Some cortical landmarks are indicated in Fig. 53 (External lanfmarks). The central sulcus (sulcus of Rolando) and the lateral sulcus (sylvian fissure) demarcate lobes of the cerebral cortex, which are named for the overlying bones of the skull: frontal, parietal and temporal. The smaller occipital lobe forms the posterior pole of the hemisphere, and the insula (or insular lobe) is the cortex of the expanded floor of the lateral sulcus (Fig. 57), overlying the lentiform nucleus. Areas of the cortex serve different functions, which have been determined from the effects of lesions, electrical stimulation in the course of surgery, and modern functional imaging techniques. The major functional regions are shown in Fig. 58 - Functional areas of cortex.
The thick layer of white matter separating the cerebral cortex from the corpus striatum and lateral ventricle contains bundles of fibers of three types. Association fibers connect different cortical areas of the same hemisphere. Commissural fibers connect the left and right cerebral cortices; most pass through the corpus callosum, but parts of the temporal lobes are connected by the anterior commissure. Ascending and descending fibers, connecting the cortex with subcortical regions, are known as projection fibers; the internal capsule is a site of concentration of many such fibers.
Functional pathways in the central nervous system
Two pathways are involved in conducting general sensory signals to the primary somatosensory area of the cerebral cortex: the spinothalamic system (also called the anterolateral pathway), and the medial lemniscus system (also called the dorsomedial pathway or, in clinical neurology, the posterior column system). These are summarized in Fig. 59 - Somatosensory pathways. The most obvious difference between the two systems is that the spinothalamic tracts cross the midline at segmental levels of the spinal cord, whereas the decussation of the medial lemnisci is in the caudal part of the medulla. Simple touch, temperature and pain
The spinothalamic system (and also the pathway from the spinal trigeminal nucleus shown in Fig. 59) are for the less discriminating sensations. These include the detection but not the detailed evaluation of stimuli impinging on the surfaces of skin and mucous membranes, and pressure that is sufficient to stimulate receptors in deeper tissues. Recognition of non-injurious variation in temperature (tested with warm and cool objects) is carried exclusively in this system. The spinothalamic tract is also the principal, but not the only ascending pathway conducting signals that are felt as pain. Surgical transection of the ventrolateral quadrant of the spinal cord abolishes the ability to experience pain on the opposite side of the body caudal to the lesion. If the patient survives for more than a few months the pain may return, poorly localized but often with greater severity than before. Evidently other ascending pathways can be recruited to detect painful stimuli in the absence of the spinothalamic tract.
Discriminative touch
The two-point discrimination test provides the simplest clinical assessment of the integrity of the medial lemniscus system, but transection of the dorsal column or medial lemniscus causes only a partial impairment of the detection of simultaneously touched sites. A more specific test seeks the identification of changes in orientation as well as spatial separation. This is most easily done by asking the patient to recognize a simple shape, such as a triangle or a letter, drawn on the skin with the examiner's finger or a smaller blunt instrument. The recognition of shapes requires also the integrity of the somatic sensory cortical areas of the parietal lobe. Detecting the vibration of a low frequency tuning fork applied to a bony prominence is a simple test often wrongly associated with the medial lemniscus system. Vibration can be felt when the dorsal columns have been completely transected, and the test has no localizing value in the central nervous system. Vibration is a useful and sensitive test for the integrity of large, rapidly conducting myelinated fibers in the peripheral nervous system. These are often the first to be involved in demyelinating neuropathies.
Proprioception
The conscious perception of position and movement originates principally in the muscle spindles, which are receptors that report the lengths (and also changes in lengths) of muscles. There are also receptors that detect mechanical conditions in tendons and joints, but their signals are probably used mainly for spinal reflexes and cerebellar activity, without conscious awareness. For the upper limb, conscious proprioception is mediated by the medial lemniscus system, which includes axons that ascend in the cuneate fasciculus (see Figures 54 and 59). The pathway from the lower limb is initially in the gracile fasciculus, but the axons of proprioceptive neurons end in the nucleus dorsalis (Clarke's column) of the dorsal horn (see Fig. 54). This nucleus is the source of the dorsal spinocerebellar tract, which is in the lateral white matter of the spinal cord. Clinical studies of rare lesions confined to the dorsal columns of the cervical cord reveal preservation of conscious proprioception in the lower limbs. Tracing experiments in animals indicate that branches of dorsal spinocerebellar tract axons end in group of neurons (Nucleus Z of Brodal & Pompeiano) just rostral to the gracile nucleus, which sends fibers into the contralateral medial lemniscus. This nucleus exists also in the human brain.
Other proprioceptive connections operate below the level of consciousness. They include synapses in the spinal gray matter (for stretch reflexes), the ventral spinocerebellar tract (Fig. 54) and the cuneocerebellar tract (upper limb equivalent of the dorsal spinocerebellar tract). Proprioceptive endings occur in the muscles supplied by cranial nerves; some of their central connections are noted in the next paragraph; others are outside the scope of these notes.
| Somatic sensation from the head
The
skin of the posterior half of the human head is innervated by the
ganglion of nerve C2. (C1 does not have a dorsal root.)
|
The primary sensory neurons that innervate muscle spindles have axonal branches in the spinal cord (or brain stem) that form excitatory synapses with motor neurons. Rapid stretching of a muscle (by tapping its tendon) evokes a monosynaptic reflex contraction: the stretch reflex or tendon jerk. Ordinary movements do not elicit this reflex contraction, because it is suppressed by activity in tracts that descend from the brain stem and cerebral cortex. Below the level of a complete transection of the spinal cord, after an initial period of "spinal shock," voluntary movement is impossible and the stretch reflexes are uninhibited. Every passive movement is resisted, and the muscles are in a state of tonic contraction known as spasticity. The upper and lower limbs are typically held in flexion.
Three descending tracts from the brain are principally responsible for modulating spinal reflexes and providing instructions for skilled and unskilled movements.
The vestibulospinal tract arises from certain large neurons in the vestibular nuclei of the medulla. These cells are activated by sensed changes in position and movement of the head, and their axons end in the medial part of the ventral horn of the spinal gray matter. They stimulate contraction of the extensor muscles of the trunk and lower limb, and (in man) the flexors of the upper limb. Transection of descending motor pathways at a level rostral to the medulla causes a spastic paralysis in which the lower limbs are extended, due to unopposed action of the vestibulospinal projection.
Reticulospinal fibers arise from neurons in the medial parts of the reticular formation of the pons and medulla. Their distribution in the spinal cord, deduced from human clinico-pathological studies, is shown in Fig. 54. (Earlier notions of separate pontine and medullary reticulospinal tracts are no longer tenable.) Neurons in the reticular formation have long dendrites that are contacted by collateral branches of axons ascending and descending through the brain stem. For example, many spinothalamic tract fibers have branches that synapse with reticular formation neurons, and there are also spinoreticular fibers. The motor regions of the reticular formation also receive descending afferents from all the motor areas of the cerebral cortex, providing a disynaptic cortico-reticulo-spinal pathway.
Corticoreticular fibers probably do not cross the midline, and most reticulospinal fibers probably decussate in either the brain stem or the spinal cord (Fig. 60 - Descending motor pathways).
The disabling "upper motor neuron" spastic hemiplegia that follows destruction of the motor and premotor cortical areas, or transection of descending motor fibers by a lesion in the internal capsule or cerebral peduncle, is probably attributable to loss of the corticoreticular projection. Transection of reticulospinal fibers probably accounts for spasticity due to destructive lesions in the spinal cord or ventral medulla.
The corticospinal tract contains the axons of cells in the primary motor area, premotor area and supplementary motor area of the frontal lobe. (The corticospinal tract also contains many fibers from the parietal lobe, but these end in the dorsal horn and probably do not serve a motor function.) Corticobulbar fibers have similar origins, but end in and around the motor nuclei of cranial nerves V, VII and IX-XII in the pons and medulla. The premotor and supplementary motor areas (see Fig. 58) also send association fibers to the primary motor cortex. All the motor areas of the cerebral cortex receive indirect input from the basal ganglia and cerebellum (Fig. 61 - Plan of motor connections in brain).
Descending pathways to the motor nuclei of cranial nerves (including corticobulbar fibers) are both crossed and uncrossed. The only muscles controlled exclusively by the contralateral cerebral hemisphere are those of the lower half of the face (facial nerve) and the trapezius (accessory nerve). The tongue (hypoglossal nerve) is largely but not exclusively under contralateral control, and the sternocleidomastoid muscle (accessory nerve) is controlled by the ipsilateral cerebral hemisphere. All other muscles of the head and pharynx, including the upper half of the face, are controlled by both cerebral hemispheres. Consequently, the paralysis due to a lesion in a cerebral hemisphere, such as infarction of the internal capsule or frontal lobe, involves only the lower half of the contralateral side of the face. A facial weakness that also involves the muscles around and above the eye can be due only to a "lower motor neuron" lesion, involving either the facial motor nucleus or its efferent axons in the brain stem or facial nerve.
The eye muscles are not directly controlled by the cerebral cortex, and will be discussed later.
Corticospinal and corticobulbar fibers, accompanied by corticoreticular fibers and corticopontine fibers (which end in the pontine nuclei), descend through the posterior limb of the internal capsule, between the thalamus and caudate nucleus (shown in Fig. 57), and then pass into the ventral part of the cerebral peduncle (see Fig. 55). At this level, the corticospinal and corticobulbar fibers are flanked medially and laterally by corticopontine fibers. The location of the corticoreticular fibers in the human midbrain has yet to be discovered. In the ventral pons, the corticospinal tracts form several small fasciculi dispersed among the pontine nuclei and the decussating pontocerebellar fibers. At the caudal border of the pons, the corticospinal fibers reassemble on each side to become the left and right pyramids of the medulla (Figures 53 and 55). This anatomical landmark accounts for a much misused synonym, pyramidal tract. At the caudal end of the medulla, most of the axons in the pyramids decussate and enter the lateral white matter of the spinal cord (see Fig. 54). Corticospinal axons end by contacting both interneurons and primary motor neurons in the ventral horn.
A small ventral corticospinal tract in the medial part of the ventral spinal white matter, consists of fibers that did not decussate in the medulla. Most of these axons cross the midline in the cord and end in the medial part of the ventral horn. A few end ipsilaterally, providing for control of the axial musculature by both cerebral hemispheres. Rarely, the ventral corticospinal tract is large with most of its fibers crossing at various segmental levels, and causing diagnostic difficulties with unilateral spinal lesions.
Physiological studies in monkeys and clinicopathological investigations of certain rare human lesions indicate that the corticospinal tracts are essential for skilled movements of the hands. Selective transection (possible only in the midbrain or medullary pyramid) causes a transient flaccid hemiplegia, followed by recovery of all voluntary movements except those requiring collaboration of different fingers. Before about 1955 it was widely believed that the corticospinal tracts mediated most or all of the supraspinal modification of spinal reflexes, and the adjective "pyramidal" was applied to syndromes of spastic paralysis resulting from large destructive lesions in the cerebral cortex or internal capsule. This misguided usage of words is still occasionally seen in clinical case reports. The term "upper motor neuron paralysis" is more appropriate for the condition in which muscles are hypertonic, with exaggerated stretch reflexes, and cannot be voluntarily used because of transection of descending motor pathways.
A traditional and still valid test of the integrity of the corticospinal or pyramidal tract is the plantar reflex. The normal response to a possibly injurious stimulus (plantar flexion) is replaced by a more primitive withdrawal (extension of the hallux, with flexion at the knee and hip: the Babinski reflex).
Other circuits for movement
The motor areas of the cerebral cortex cannot control the skeletal musculature without assistance from the cerebellum, which looks after the timing and duration of volleys of impulses in motor neurons, and the basal ganglia, which provide patterns of neuronal activity for learned skills, which are complicated but not directly willed at the time the movements are made.
Cerebellar circuits
The connections of the cerebellum have already been described. In Figure 61 they are placed in context with descending pathways from the cerebral cortex and the circuitry of the basal ganglia.
Basal ganglia circuits
The expression "basal ganglia" embraces the corpus striatum (caudate and lentiform nuclei, shown in Fig. 57), the substantia nigra in the midbrain, and the subthalamic nucleus (see Fig. 56). The lentiform nucleus comprises adjacent but functionally different parts. The putamen and the caudate nucleus constitute the striatum. The medial part of the lentiform nucleus (globus pallidus, shown in Fig. 57) is the pallidum, which has differently connected external and internal divisions. The circuitry and neurotransmitters of the basal ganglia are quite well understood (Fig. 62 - Basal ganglia circuitry), and this knowledge provides a crude understanding of some disorders of the system. For example, destruction of the subthalamic nucleus leads to reduced inhibition of the thalamus, which then stimulates the motor cortical areas excessively, causing contralateral hemiballismus. A similar argument may account for the poverty of movement in Parkinson's disease, which is due to degeneration of the dopamine-producing neurons of the substantia nigra.
Eye movements
The neuronal circuits that control movement of the eyes are numerous and complicated, being driven largely by the sensory visual system. Some of these connections are shown in Fig. 63 - Eye movement connections. Salient features of this system are (1) For vertical eye movements, the pathway involves nuclei in the rostral midbrain, and (2) For horizontal eye movements the pathway descends to the caudal pons (abducens nucleus, for lateral rectus muscle), then crosses the midline and ascends to the rostral midbrain (oculomotor subnucleus for medial rectus muscle). Convergence and divergence of the eyes are regulated mainly at the level of the midbrain and do not require the integrity of the frontal eye field.
At least two cortical areas are involved in the control of eye movements. The frontal eye field (Figs 58, 63) stimulates rapid shifting of the direction of gaze to the opposite side. Destruction of this cortical area results in a conjugate deviation of the eyes toward the side of the lesion. Pursuit movements (tracking an object moving slowly across the field of vision) are not paralyzed. The parieto-occipital eye field, which is coextensive with much of the visual association cortex, is the region needed for pursuit. Lesions here also cause visual defects but do not prevent rapid, voluntary eye movements.
Special senses
Equilibration
The static vestibular receptors in the utricle and saccule of the inner ear respond to the position of the head in relation to gravity (or other accelerating or decelerating forces), and the kinetic receptors in the semicircular ducts report rotational movement of the head in any plane. The primary sensory neurons are in the vestibular ganglion, and their axons enter the brain in the vestibular division of cranial nerve VIII (Table 7 - Cranial nerve components). Most end in the four vestibular nuclei, which are beneath the lateral part of the floor of the fourth ventricle; some go directly to the cerebellum in its inferior peduncle. The reciprocal connections of the vestibular nuclei with the vestibular parts of the cerebellum were discussed in the context of the cerebellum, and the vestibulospinal tract was described with the other descending motor tracts (Figs 60, 61).
A poorly understood ascending pathway from the vestibular nuclei to the thalamus and then to the cerebral cortex provides conscious awareness of position and movement as perceived by the receptors in the inner ear. Cortical areas responding to stimulation of the kinetic receptors have been identified at various sites in the parietal and temporal lobes. The principal symptom of abnormal stimulation or inhibition of the vestibular system is vertigo associated with nystagmus (see below) and nausea or vomiting. The gastric disturbance is attributed to disrupted connections of the vestibular nuclei with one of the preganglionic parasympathetic nuclei of the vagus nerve. A unilateral destructive lesion in the inner ear, vestibular nuclei or inferior cerebellar peduncle also causes ataxia, with a tendency to fall toward the side of the lesion. This is attributed to unopposed action of the contralateral vestibulospinal tract on motor neurons that supply extensor muscles of the lower limb.
Some neurons in the vestibular nuclei have axons that travel rostrally in the medial longitudinal fasciculus (MLF), both ipsilaterally and contralaterally, and end in the motor nuclei of cranial nerves VI, IV and III. The MLF is adjacent to the midline at all levels of the brain stem. It contains also the axons of neurons used for rapid conjugate eye movements (Fig. 63).
Vestibulo-oculomotor fibers in the MLF mediate the vestibulo-ocular reflex, which is a slow conjugate movement of the eyes in a direction opposite to that of a slow rotation of the head through a small angle. The angular movement of the eyes is equal to that experienced by the semicircular ducts, but in the opposite direction. This reflex is disturbed by rapid movements of endolymph in the semicircular ducts, which cause vertigo and back-and-forth eye movements known as nystagmus. When nystagmus is due to vestibular stimulation, each cycle of eye movement has a slow component in one direction (driven by the vestibular system) and a fast corrective component (driven by descending pathways; see Fig. 63). In caloric testing the kinetic receptors are artificially stimulated by irrigating the external ear with warm or cool water, causing convection currents in the semicircular ducts. The normal response is nystagmus with slow and fast components. In a comatose patient, only the slow component is seen because the fast "voluntary" compensation is suppressed. The eyes move toward the side stimulated by cool water. Absence of this response after bilateral stimulation indicates destruction of the vestibular nuclei, medial longitudinal fasciculi or ocular motor nuclei, and contributes to a diagnosis of brain stem death.
Hearing
Neurons in the spiral ganglion of the cochlea send their axons to the cochlear nuclei, which are located on the dorsolateral and ventrolateral aspects of the inferior cerebellar peduncle. The afferents and efferents of the cochlear nuclei are organized according to the frequencies of sounds received by the organ of Corti in the inner ear. The cochlear nuclei project bilaterally to other nuclei in the brain stem, and the pathway ascends bilaterally, with relays in the midbrain and thalamus, to the primary auditory area on the superior surface of the temporal lobe. The auditory association cortex, necessary for recognition and interpretation of sounds is posterior to the primary area, and in the left hemisphere it is coextensive with part of the receptive language area (Fig. 58).
Unilateral deafness does not occur with transection of the auditory pathway rostral to the cochlear nuclei (rostral medulla), and is therefore attributable to disease of the ear (common) or lesions that impinge on cranial nerve VIII or the inferior cerebellar peduncle (rare). Obstruction of the anterior inferior cerebellar artery can cause unilateral deafness, associated at first with vertigo.
Vision and visual reflexes
The visual pathway
There is a topographical projection of the visual field throughout the pathway from the retina to the primary visual cortex, which is in and adjacent to the calcarine sulcus (see Fig. 58). Partial decussation in the optic chiasma ensures that axons from the medial half of each retina cross the midline and project to the contralateral cerebral hemisphere, whereas the lateral half of the retina projects ipsilaterally. This arrangement ensures that for each eye signals from the left or right visual field are sent to the contralateral thalamus and cerebral cortex (see Fig. 64 - Human visual pathway).
A unilateral lesion that interrupts the visual pathway posterior to the optic chiasma will cause blindness in the contralateral visual fields of both eyes. The thalamocortical fibers that loop into the temporal lobe (Meyer's loop, Fig. 64) carry signals that originated in the lower halves of the ipsilateral hemiretinas. A destructive lesion in a temporal lobe may therefore cause (among other symptoms) blindness in the contralateral upper quadrants of the visual fields. Small cortical lesions can separately involve the central and peripheral visual field. Larger lesions (such as occlusion of a posterior cerebral artery) affect the whole contralateral field, sometimes with sparing of central vision attributable possibly to an accessory blood supply to the occipital pole.
Pupillary light reflex
Not all the axons of the optic tract end in the thalamus. Some go to the superior colliculus of the midbrain, and others go to the pretectal area, which is rostral to the superior colliculus. When a bright light is shone into one eye, signals are sent into both optic tracts and to the pretectal areas of both sides. Pretectal neurons have axons that synapse with the preganglionic parasympathetic nucleus of the oculomotor nerve. Again the projection is bilateral, because some of the fibers cross in the posterior commissure, which is rostral to the pretectal area. The parasympathetic (Edinger-Westphal) nucleus sends axons to the ciliary ganglion, which is in the orbit, and the neurons in the ciliary ganglion have axons that travel in the short ciliary nerves to supply the sphincter pupillae muscle of the iris. When one retina is illuminated both pupils normally constrict. The contralateral ("consensual") response is mediated by the decussating fibers in the optic chiasma and posterior commissure. The dilator pupillae muscle is supplied by postganglionic sympathetic fibers. These are the axons of cells in the superior cervical ganglion, and they eye by way of the carotid plexus and the long ciliary nerves.
Accommodation for near vision
When the eyes converge to look at a near object, the ciliary muscle contracts, allowing the lens to thicken. This shortens the focal length of the refracting media of the eye and allows an image to be sharply focused on the retina. At the same time the pupil constricts, so that the peripheral zone of the lens, which could introduce optical aberrations, is not used. The ciliary muscle, like the sphincter pupillae, is supplied by the ciliary ganglion. The central pathway for accommodation of the lens and pupil differs, however, from that of the pupillary light reflex. Signals pass from the retinas to the superior colliculi and then to the Edinger-Westphal nucleus. The pretectal area does not form part of the pathway for accommodation. Convergence and divergence movements of the eyes are regulated by connections of the superior colliculi and other nuclei in the midbrain.
The different circuits for the light reflex and accommodation may account for the Argyll Robertson pupil. This is smaller than normal, in an eye without visual impairment, and it constricts with accommodation but not in response to light. The selective loss of the light reflex can be explained by a lesion in the pretectal area.
Smell
The primary olfactory neurons
are the receptor cells in and near the roof of the
nasal cavity. They are regularly replenished from a population of stem
cells in the epithelium, and axons of the new cells grow into the brain
and form synaptic connections there.
The unmyelinated axons of the receptor
cells gather together in the submucosal connective tissue to form about
20 olfactory nerves on each side. These nerves pass
through the small holes in the cribriform plate of the ethmoid bone,
pierce the dura and other meninges, and then spread out over the
surface of the olfactory bulb, which lies above the
cribriform plate and beneath the medial part of the orbital surface of
the frontal lobe.
Olfactory
neurosensory cells are the only mammalian neurons that can do this.
Axons in a transected and repaired dorsal root regenerate up to but not
across the peripheral-central junction. There are some parts of the
central nervous system where new neurons are generated in adult
animals. In teleost fishes the peripheral edge of the retina expands
with the growth of the eye. In adult rats it has been shown that
neurons are continually added to the dentate gyrus of the hippocampal
formation.
The primary olfactory axons enter the bulb and terminate in complex synaptic arrangements around the dendrites of large neurons known as mitral cells. Many other neurons in the olfactory bulb contribute to these synaptic complexes and contribute to the interpretation of the signals initiated by the multitude of airborne chemical stimuli. The axons of the mitral cells enter the olfactory tract, which blends with the main mass of the forebrain in the region lateral to the optic chiasma. The fibers of the olfactory tract end in (a) the uncus, which is the most medial part of the inferior surface of the temporal lobe, at the anterior end of the parahippocampal gyrus, (b) the medial part of the amygdala, which is a group of nuclei inside the temporal lobe, between the uncus and the temporal horn of the lateral ventricle, (c) a small part of the insula adjacent to the uncus, (d) the posterior part of the inferior surface of the frontal lobe, on the opposite side of the lateral sulcus from the uncus, and (e) in the entorhinal area, which is the anterior end of the parahippocampal gyrus. The entorhinal area also receives association fibers from olfactory areas (a) to (d), and it has important connections with the hippocampus and other parts of the brain involved in memory.
Loss of the sense of smell (anosmia) is usually due to disease in the nose, but it can also follow head injury, with transection of the olfactory nerves at the level of the cribriform plate. Excitatory lesions in the medial part of the temporal lobe cause olfactory hallucinations and form part of the aura of uncinate attacks, a form of temporal lobe epilepsy named from the early involvement of the uncus.
Taste
The receptor cells of taste buds have synapse-like connections with neurons whose cell bodies are in the sensory ganglia of cranial nerves VII, IX and X (Table 7 - Cranial nerve components). The central projection is to the gustatory nucleus. This is the rostral part of the solitary nucleus, a column of cells that extends through the length of the medulla. The neurons in the gustatory nucleus have axons that ascend ipsilaterally in the brain stem; they end in a thalamic nucleus that projects to a cortical area below and just behind the lower end of the primary somatic sensory area (Fig. 58). This gustatory cortex extends onto the insula, which is a cortical region involved in olfactory, gustatory and autonomic functions.
Loss of taste sensation (ageusia) is not a common complaint but it can occur in middle ear disease, because the chorda tympani nerve carries taste fibers that serve the anterior third of the tongue. Sweet tastes are not perceived, but bitter tastes (detected by receptors on the posterior part of the tongue) are unimpaired. Damage to the facial nerve in the subarachnoid space (as from an acoustic neuroma) also impairs taste sensations from the soft palate, which is supplied by the most proximal branch of the facial nerve.
Language, memory and behavior
The most complex activities of the brain are carried out in the cerebral cortex and in certain connected subcortical structures.
Language and speech
The recognition and interpretation of spoken and written language occur unilaterally, in the left cerebral hemisphere of most people. Association and commissural fibers connect the auditory and visual areas with the posterior part of the superior temporal gyrus (Wernicke's area) and the nearby cortex of the parietal lobe (angular gyrus). Destructive lesions in this receptive language area result in receptive aphasia: inability to understand speech and writing. This is associated with the production of garbled speech, which also is not understood by the speaker. The production of speech requires the integrity of an expressive speech area (Broca's area) in the frontal lobe (Fig. 58), on the same side as the receptive area. Damage to Broca's area leads to expressive aphasia, in which the attempt to speak results in an output of meaningless, jumbled words. These are heard and recognized as nonsense by the patient.
Cortical areas in the hemisphere not dominant for language are involved in prosody, a combination of tones and emphasis that has emotional and musical content, without which the voice is monotonous. The right hemisphere is also used for spatial awareness, including recognition of parts of the body, and for the recognition of unseen objects held in the hand.
Memory
The formation of new memories occurs in the circuitry of the hippocampus, which is inside the temporal lobe. Association and commissural fibers connect all the sensory association areas with the entorhinal area (see Fig. 58), and this cortex projects to the hippocampus. Efferent fibers of the hippocampus travel in the fornix to the mamillary body of the hypothalamus, which sends axons to the anterior nuclei of the thalamus. This nucleus projects to the anterior part of the cingulate gyrus, on the medial surface of the hemisphere, which is connected by subcortical association fibers with the entorhinal area. The cingulate gyrus, entorhinal area and intervening cortex constitute the limbic lobe (Fig. 58), and the hippocampus and connected parts of the brain form part of a "limbic system," together with the amygdala and certain nuclei in the brain stem.
Bilateral interruption of hippocampal circuitry described above results in loss of the ability to form new memories. Older memories can still be recalled, probably because they are stored diffusely throughout the cerebral cortex. Lesions may be in the entorhinal area and hippocampus (Alzheimer's disease), the fornices (damaged by tumors or surgery) or the mamillothalamic tracts (Korsakoff's psychosis). The hippocampi or fornices may also be damaged by separate vascular occlusions, usually of branches of the posterior cerebral arteries.
Amygdala and prefrontal cortex
The amygdala is a group of nuclei in the anterior part of the temporal lobe. Its olfactory afferents have already been mentioned. Other nuclei in the amygdala have extensive two-way connections with the prefrontal cortex (the frontal pole and the orbital surface of the frontal lobe), and with the cortex of the temporal lobe. There are also descending connections of the amygdala with the hypothalamus, epithalamus and several nuclei of the brain stem associated with the autonomic nervous system and other involuntary functions.
Electrical stimulation of the human amygdala causes feelings of fear. Destructive lesions of the temporal lobe that involve the amygdala result in docility, flattened emotional responses and sometimes, in males, increased or perverted sexual activity.
The functions of the prefrontal cortex have been deduced largely from the consequences of brain injuries, surgery (prefrontal leukotomy) and degenerative disease (neurosyphilis, Pick's disease). Lesions must be bilateral to affect behavior. The patients become rude, inconsiderate of others, and unable to foresee the consequences of their actions. These changes in personality and behavior indicate that the prefrontal cortex is involved in the planning of complex activities and making of decisions.
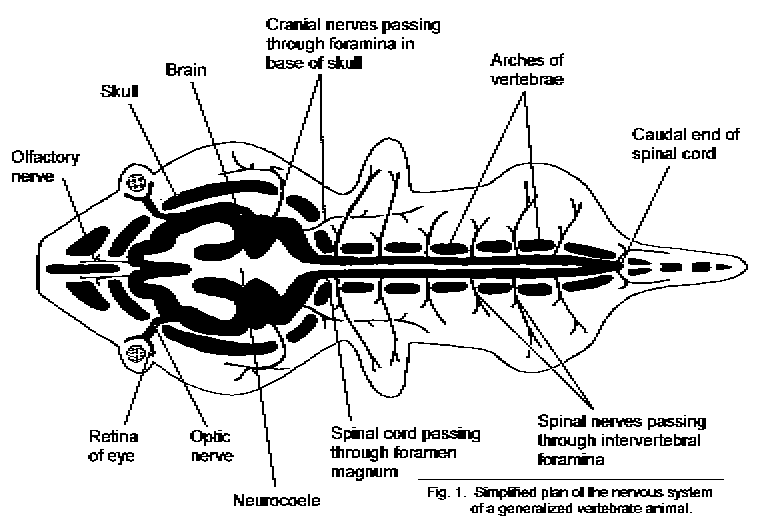{kind=link}
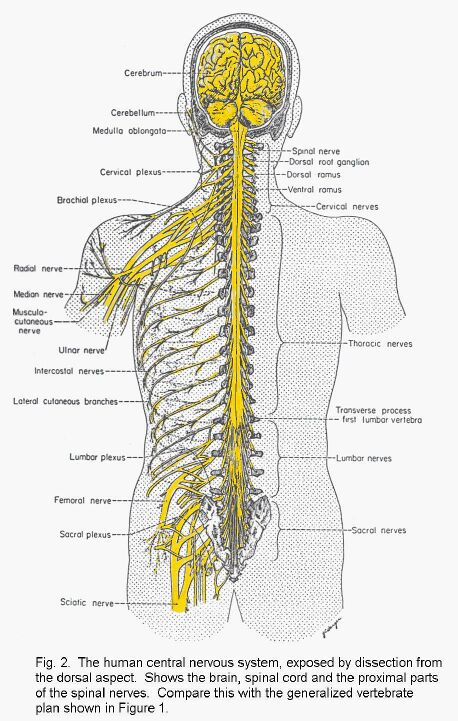{kind=link}
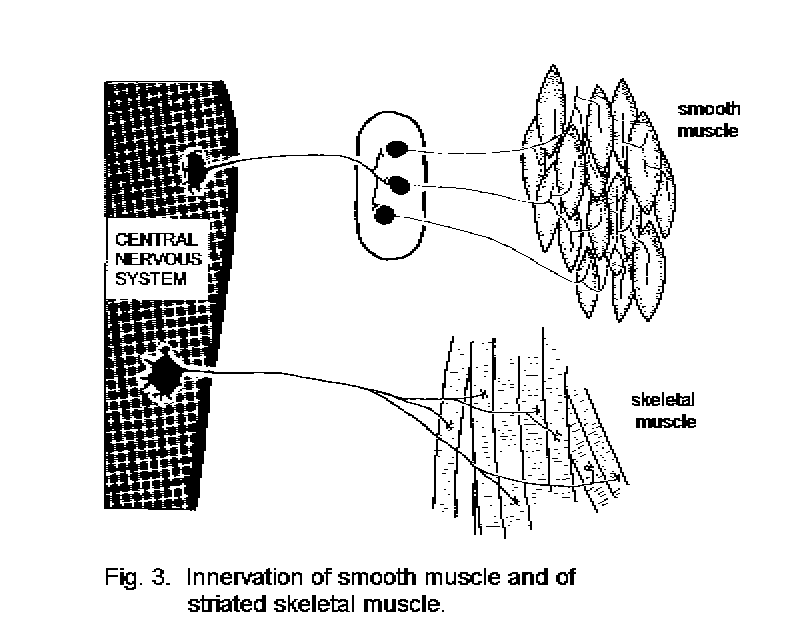{kind=link}
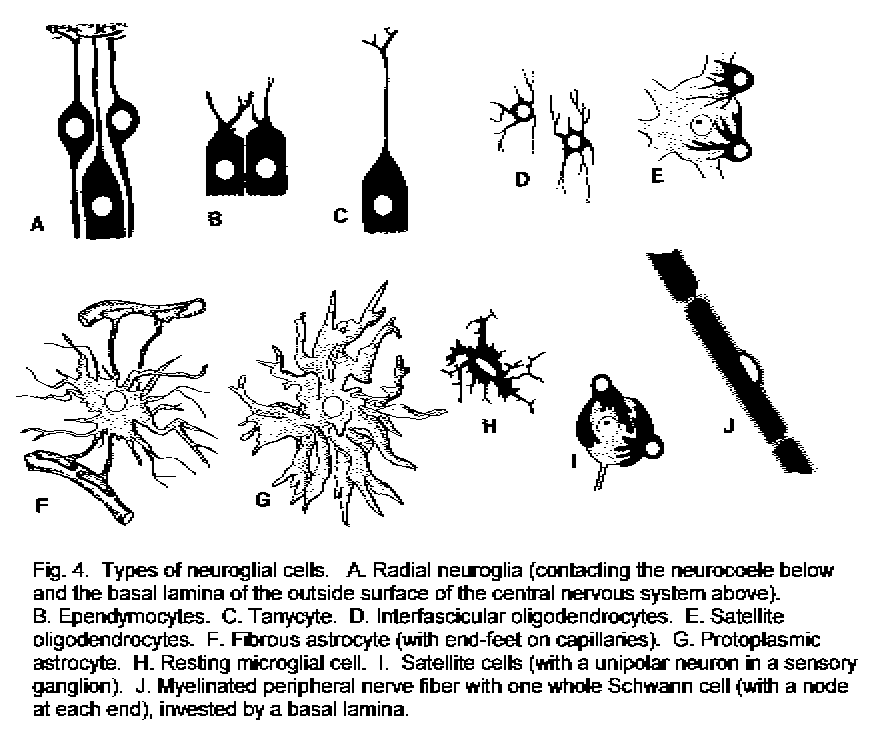{kind=link}
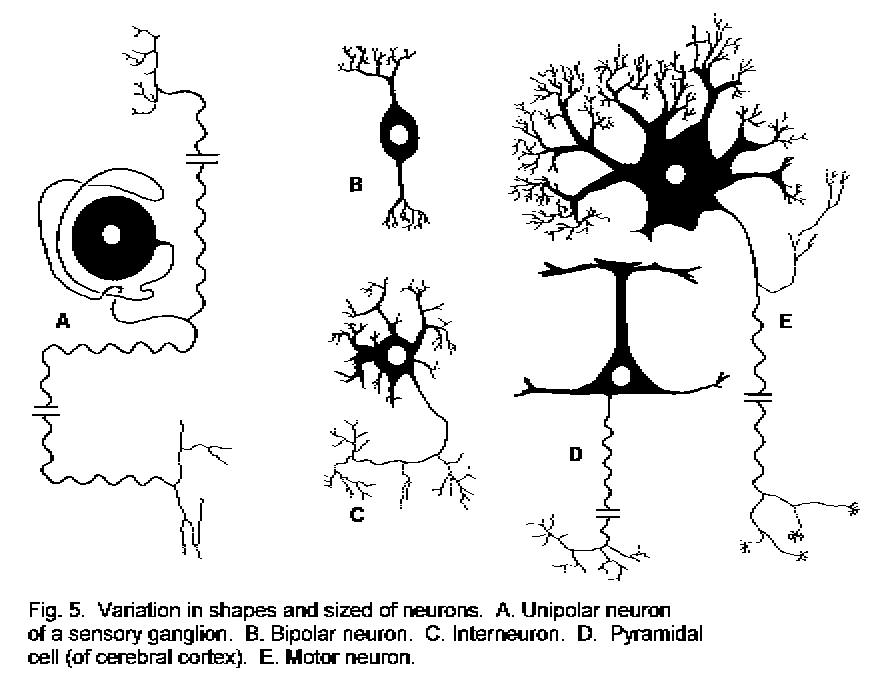{kind=link}
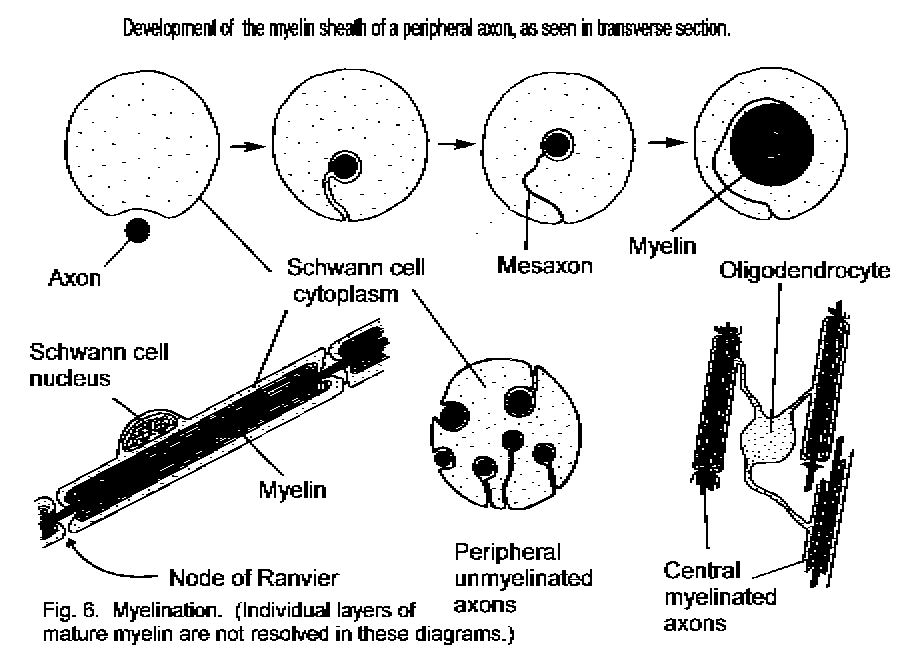{kind=link}
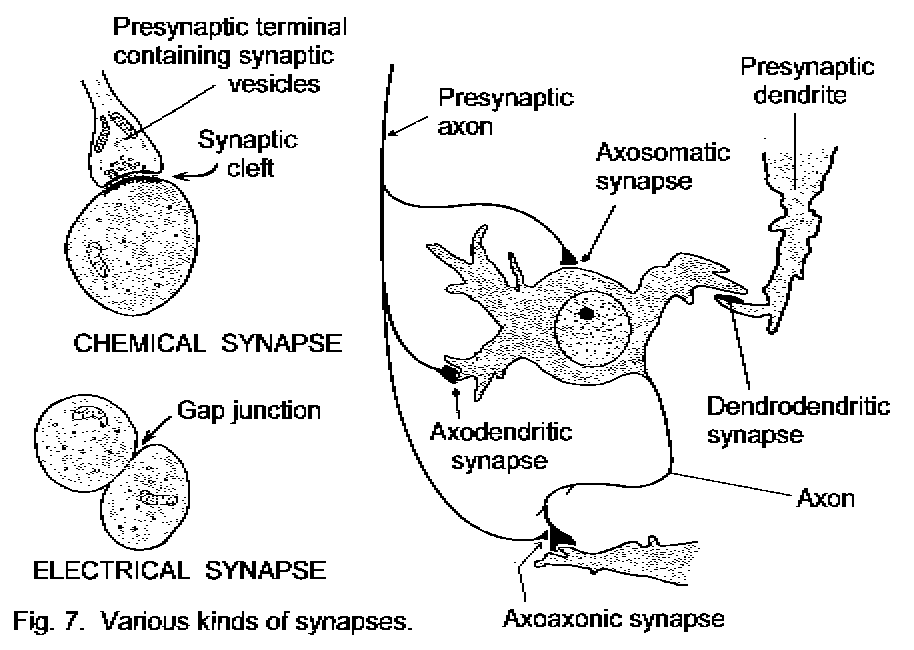{kind=link}
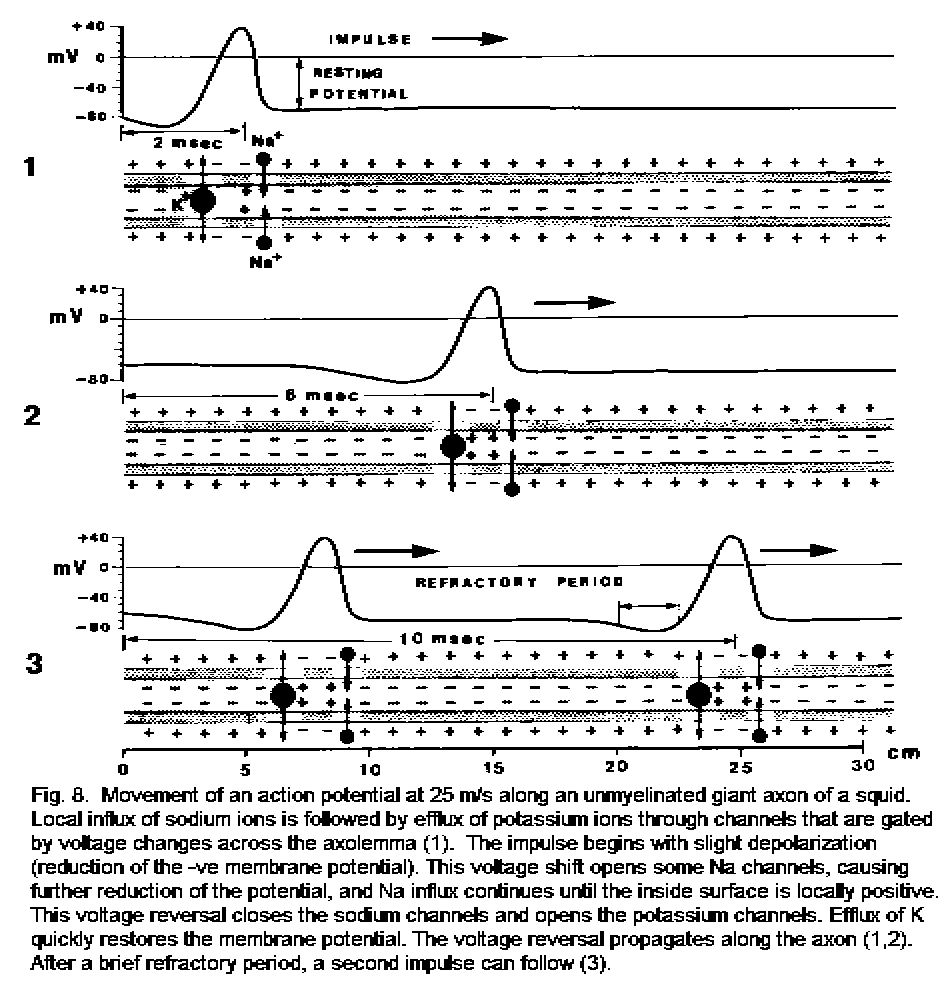{kind=link}
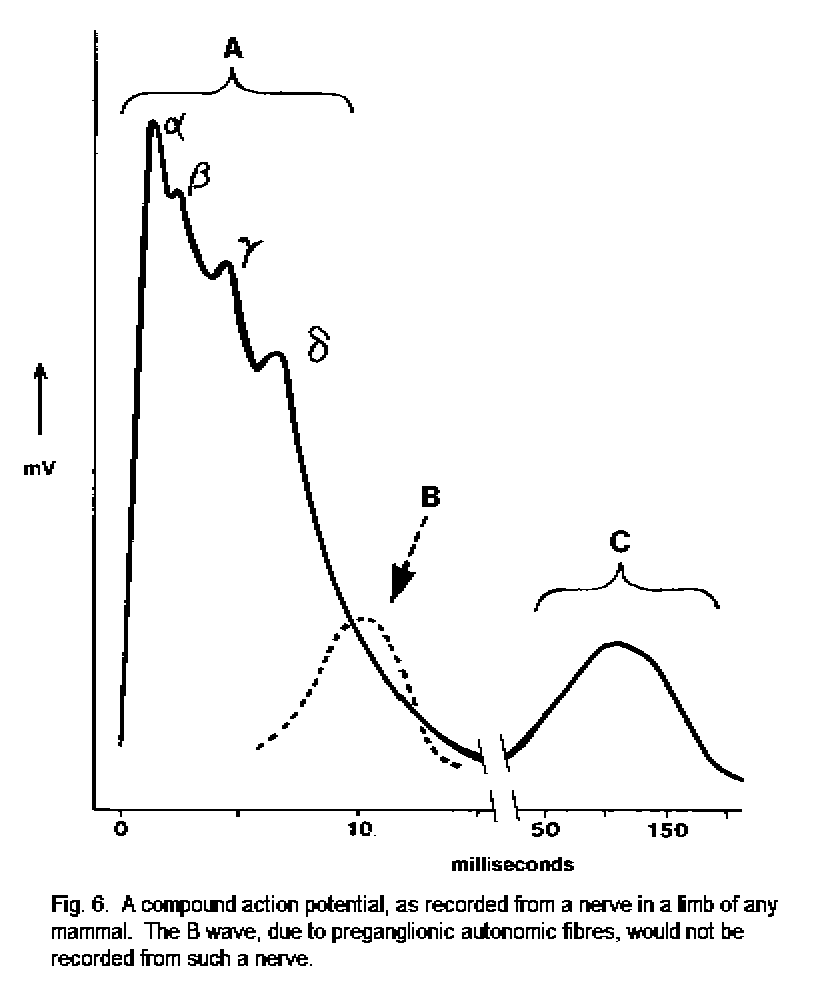{kind=link}
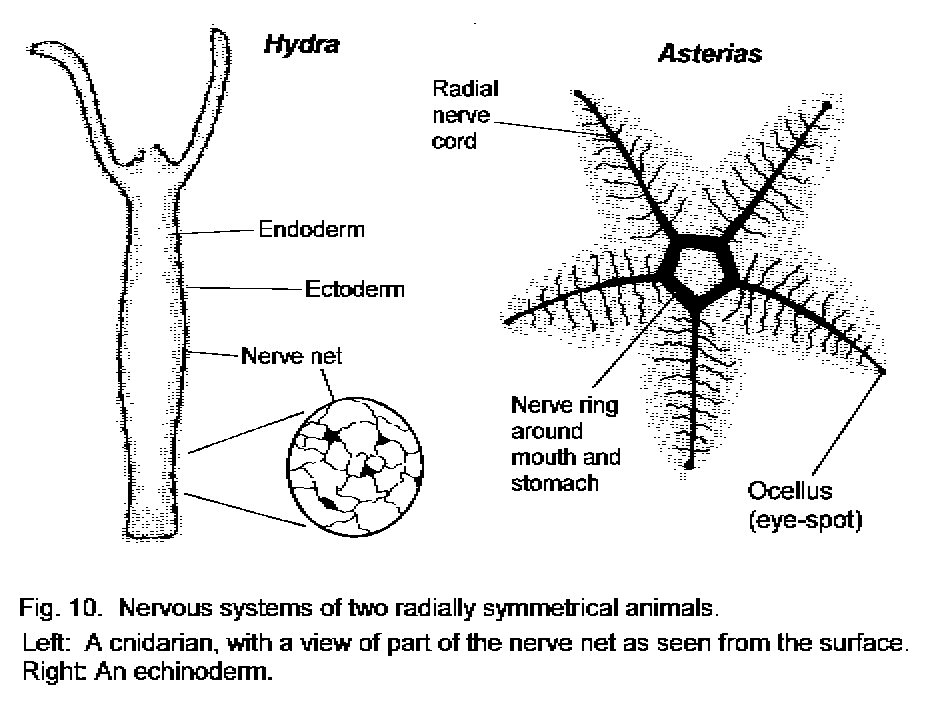{kind=link}
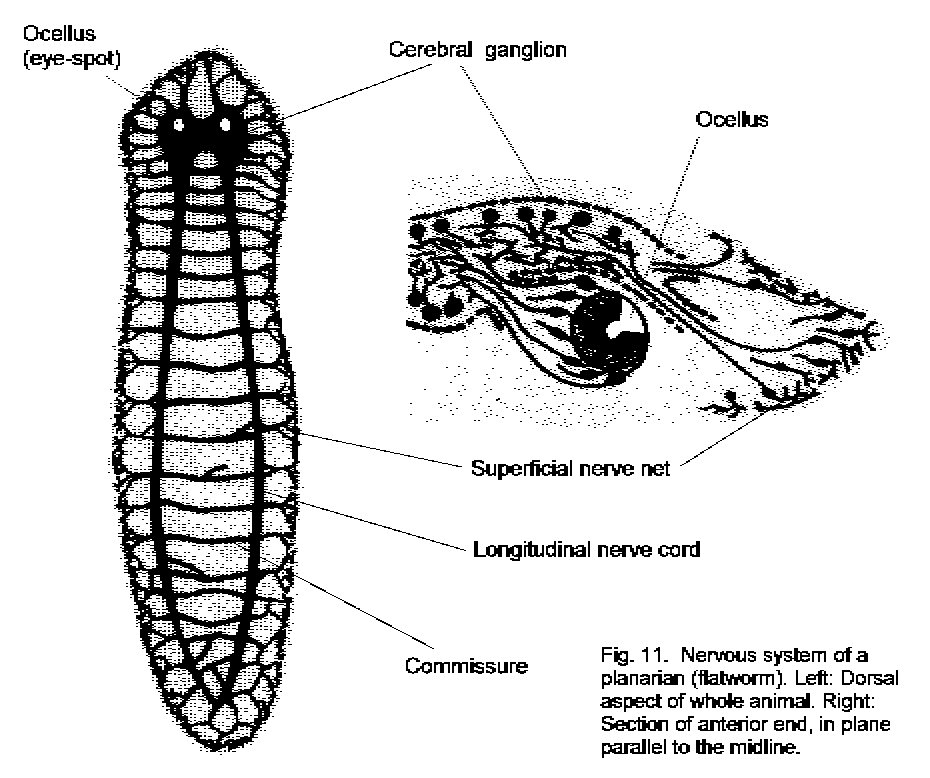{kind=link}
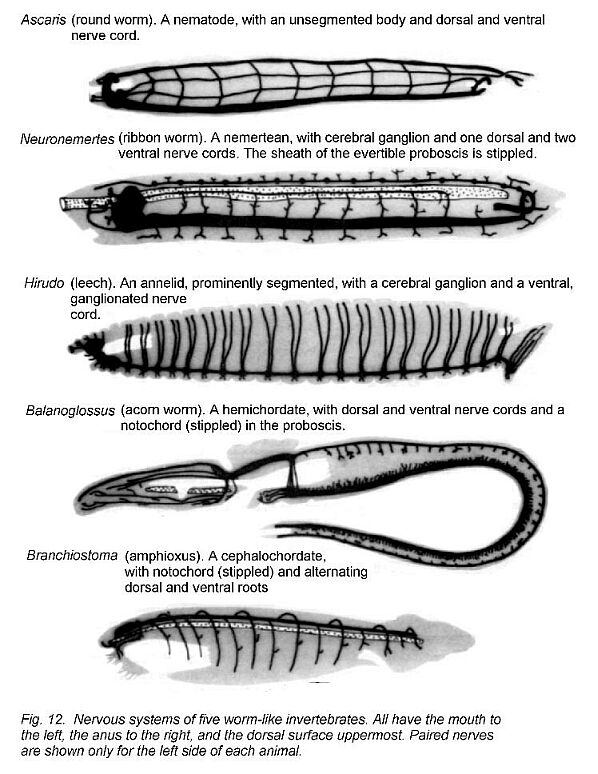{kind=link}
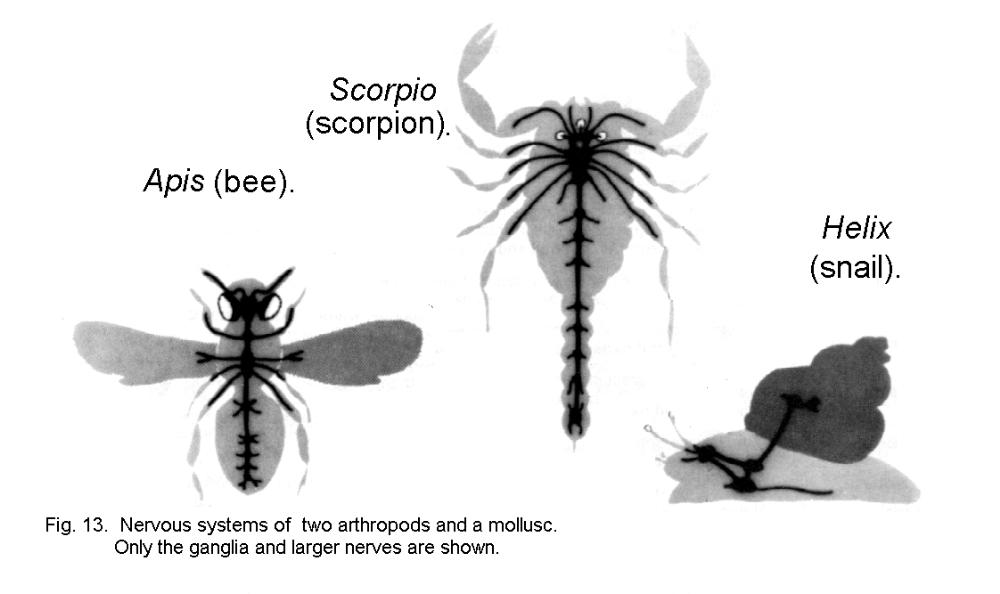{kind=link}
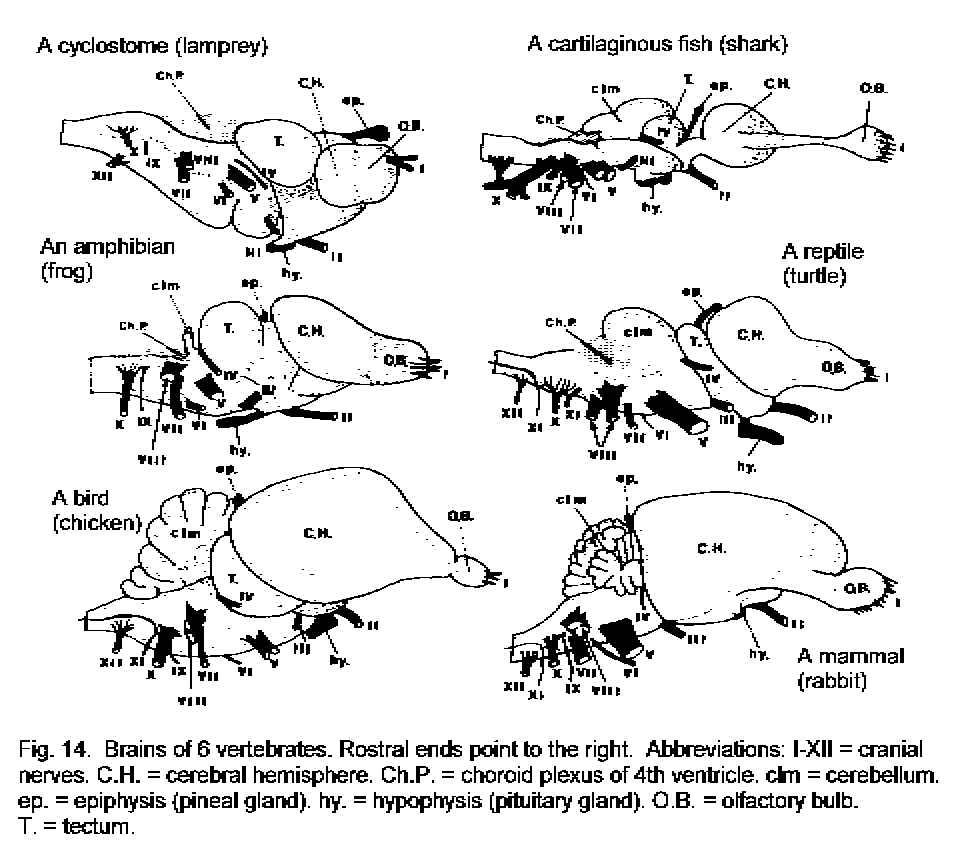{kind=link}
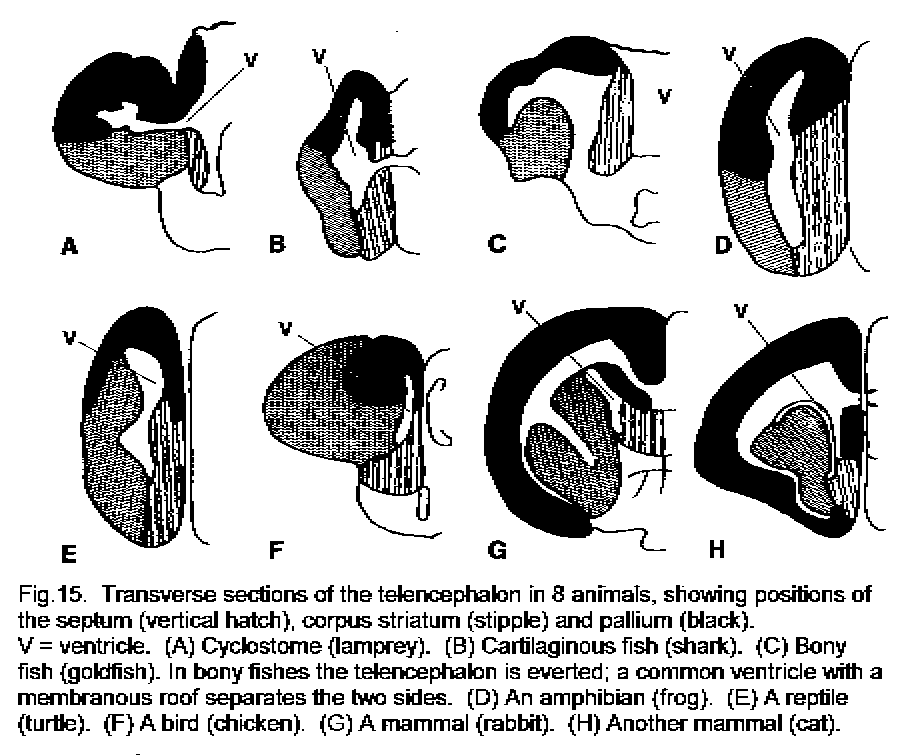{kind=link}
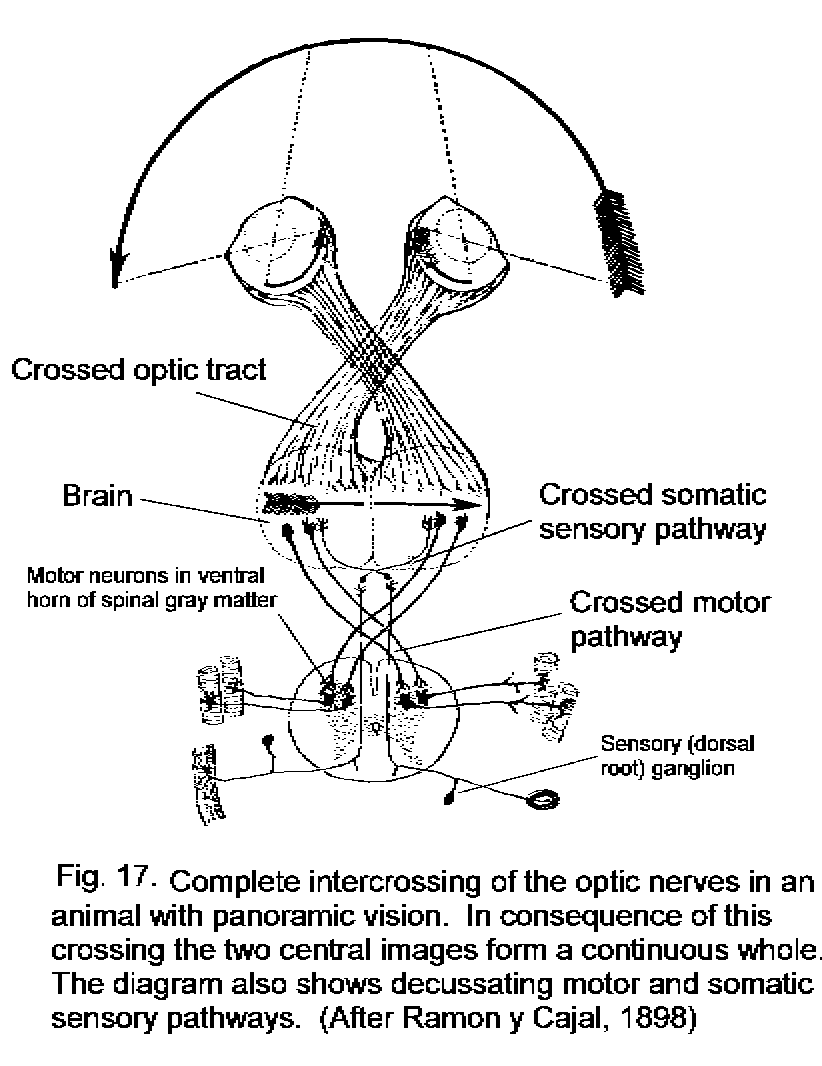{kind=link}
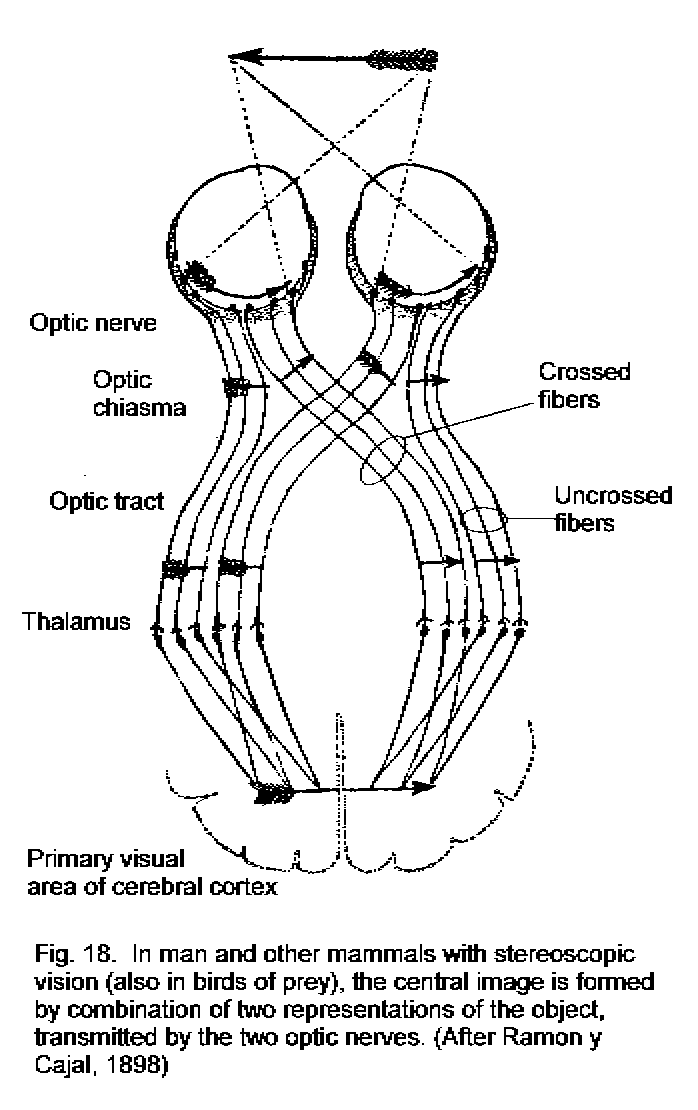{kind=link}
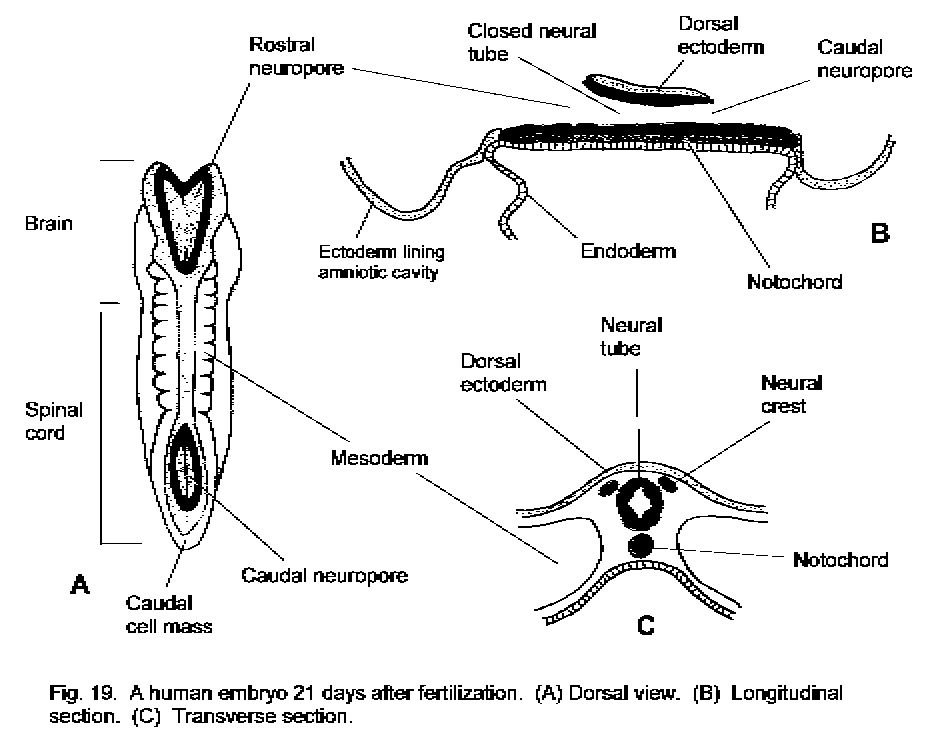{kind=link}
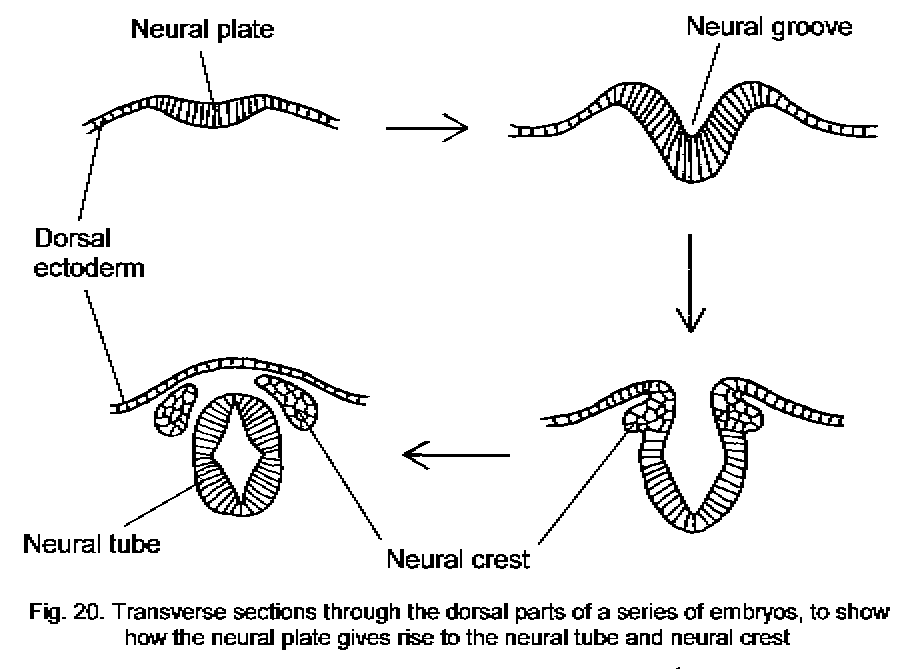{kind=link}
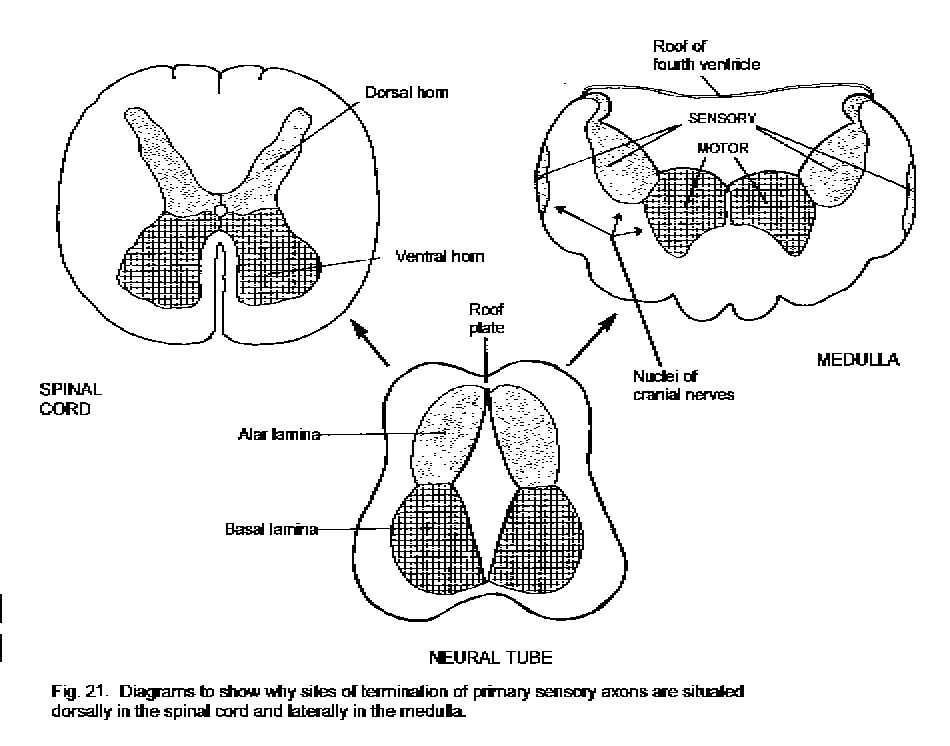{kind=link}
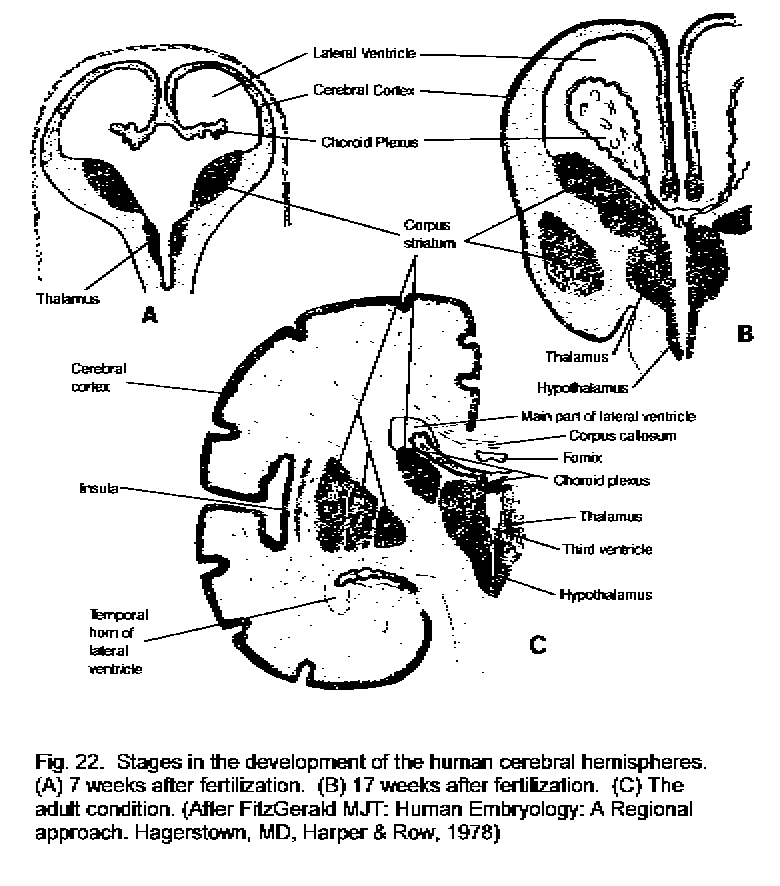{kind=link}
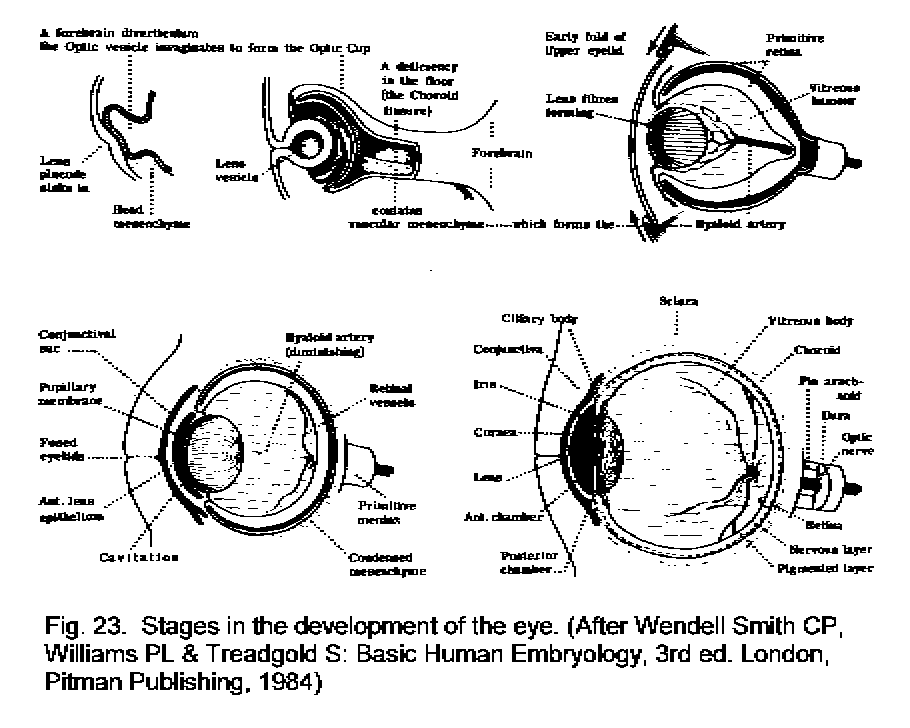{kind=link}
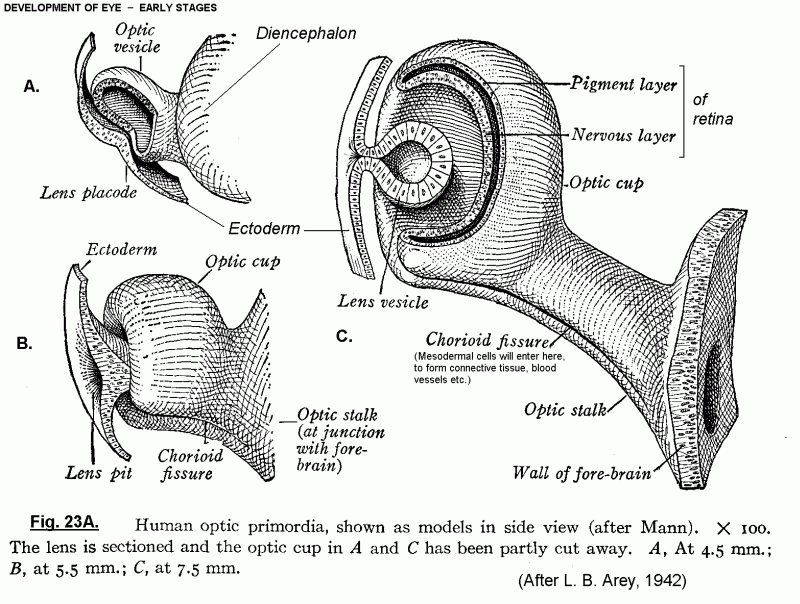{kind=link}
{kind=link}
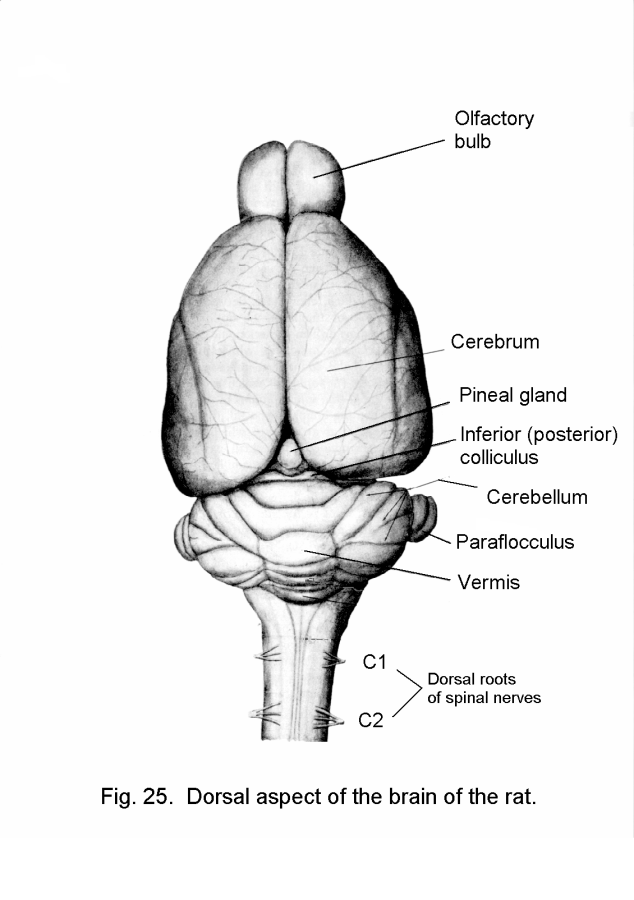{kind=link}
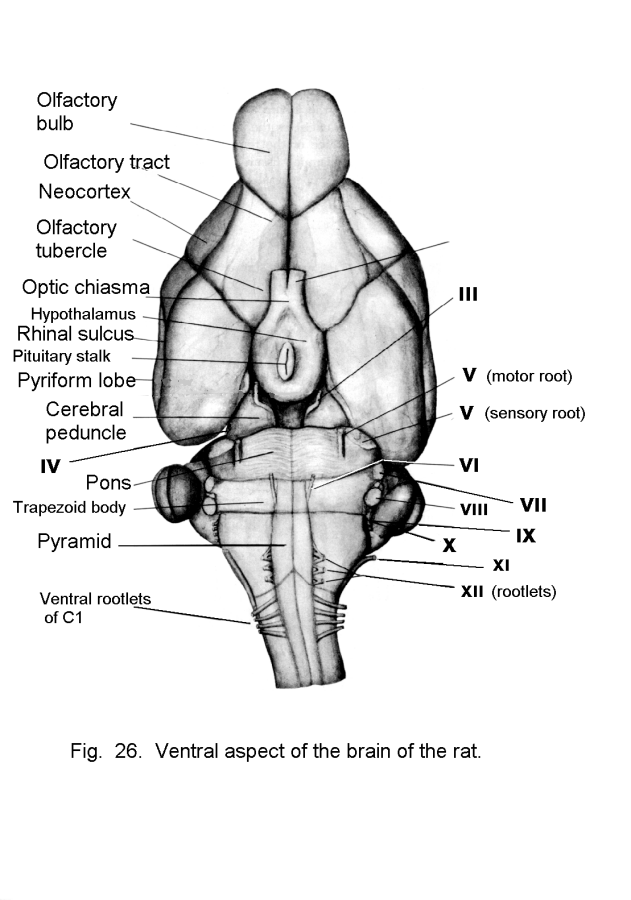{kind=link}
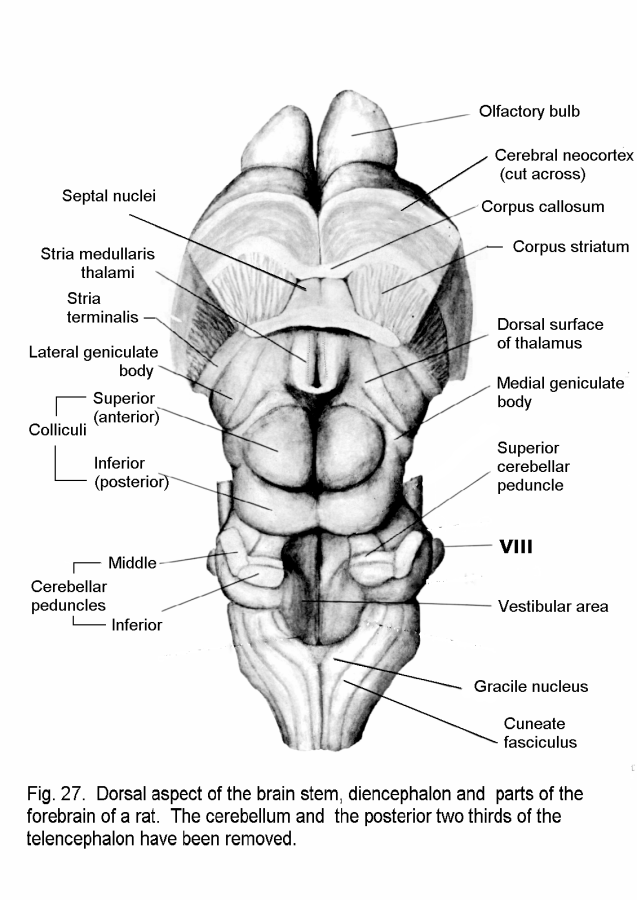{kind=link}
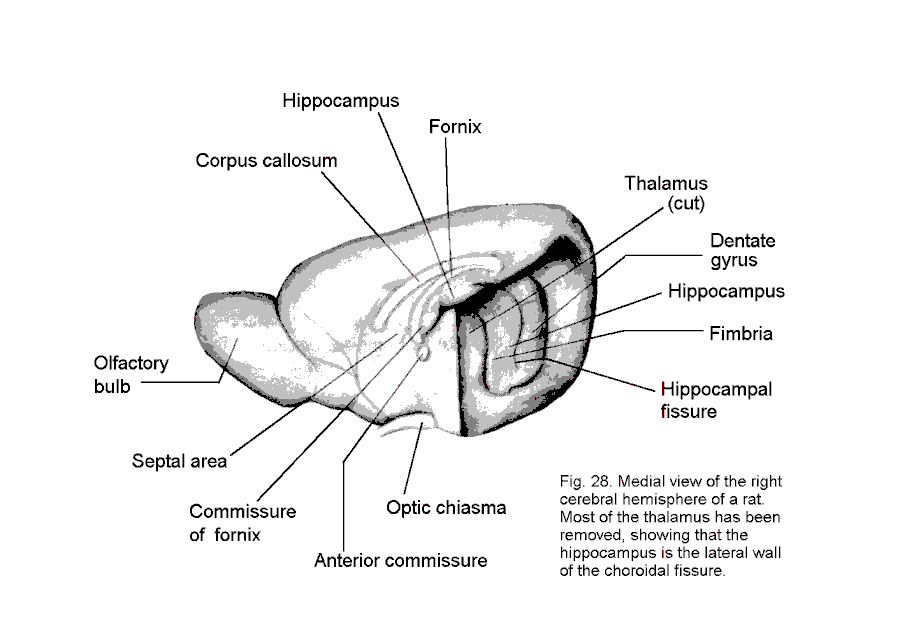{kind=link}
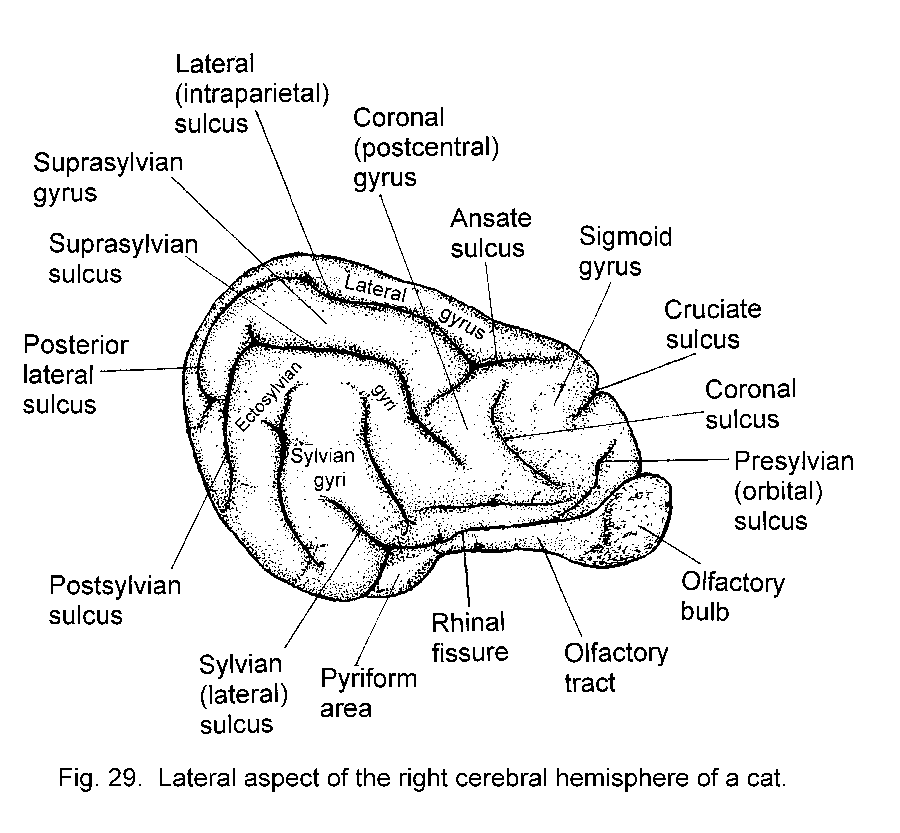{kind=link}
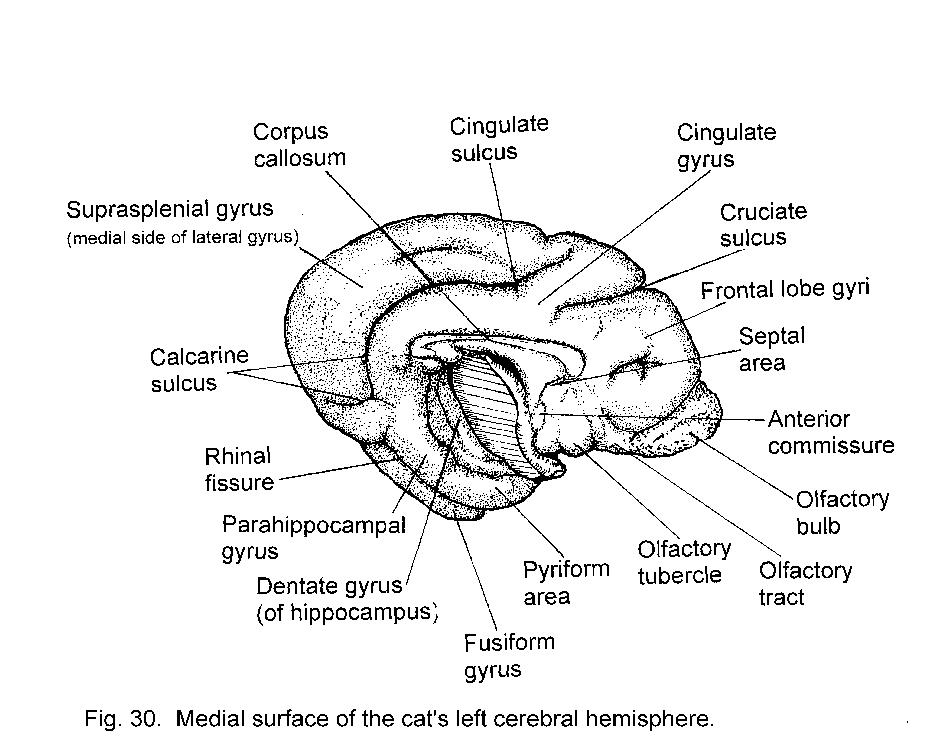{kind=link}
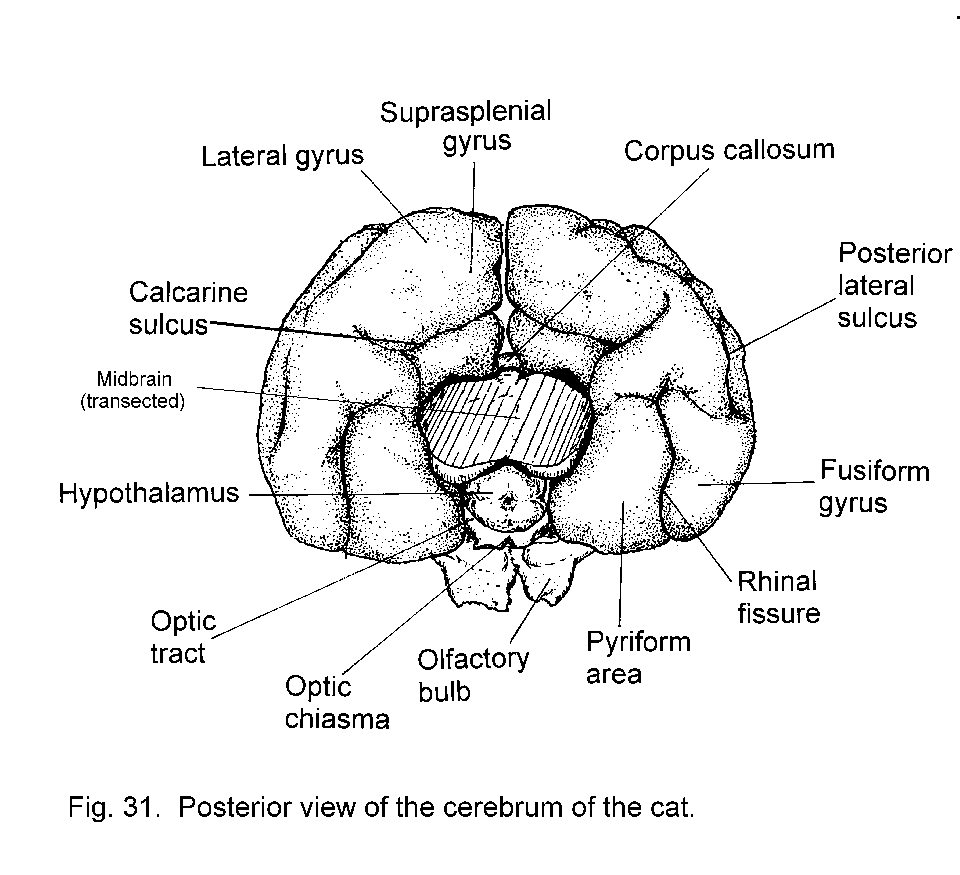{kind=link}
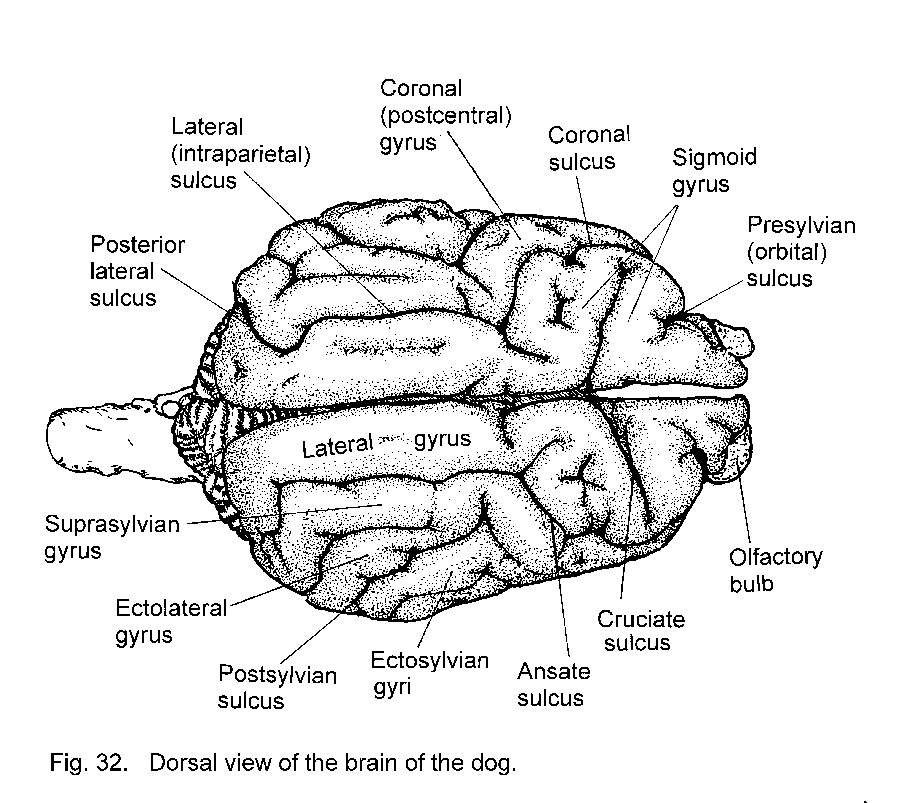{kind=link}
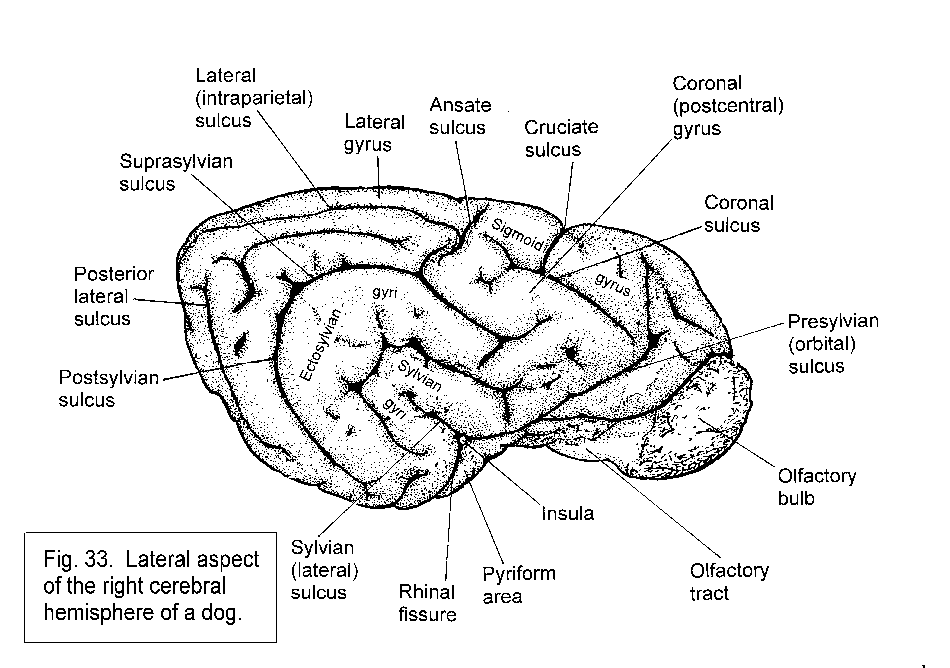{kind=link}
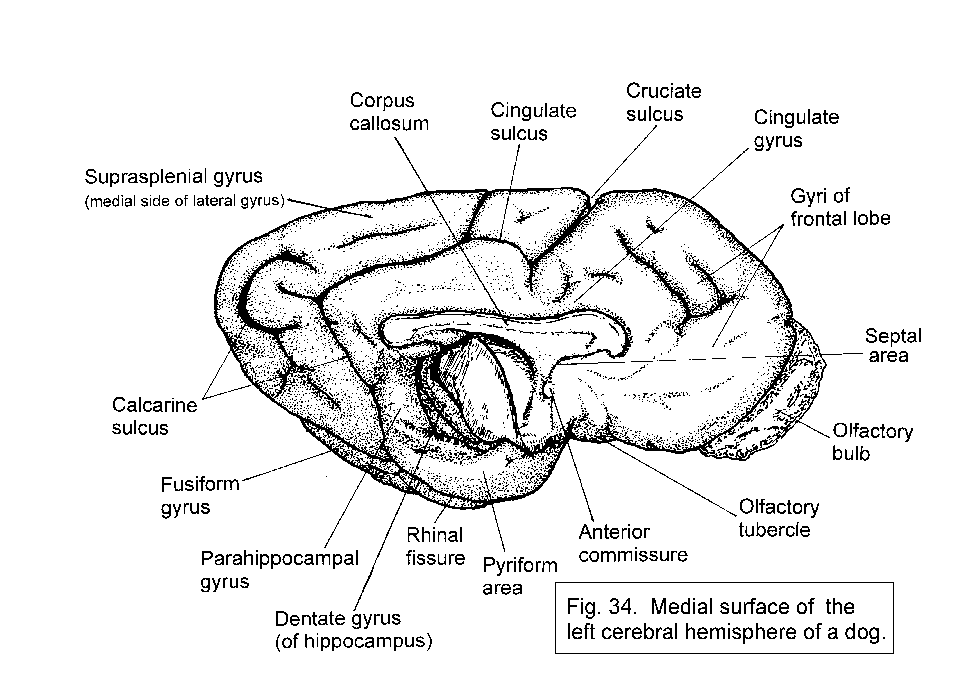{kind=link}
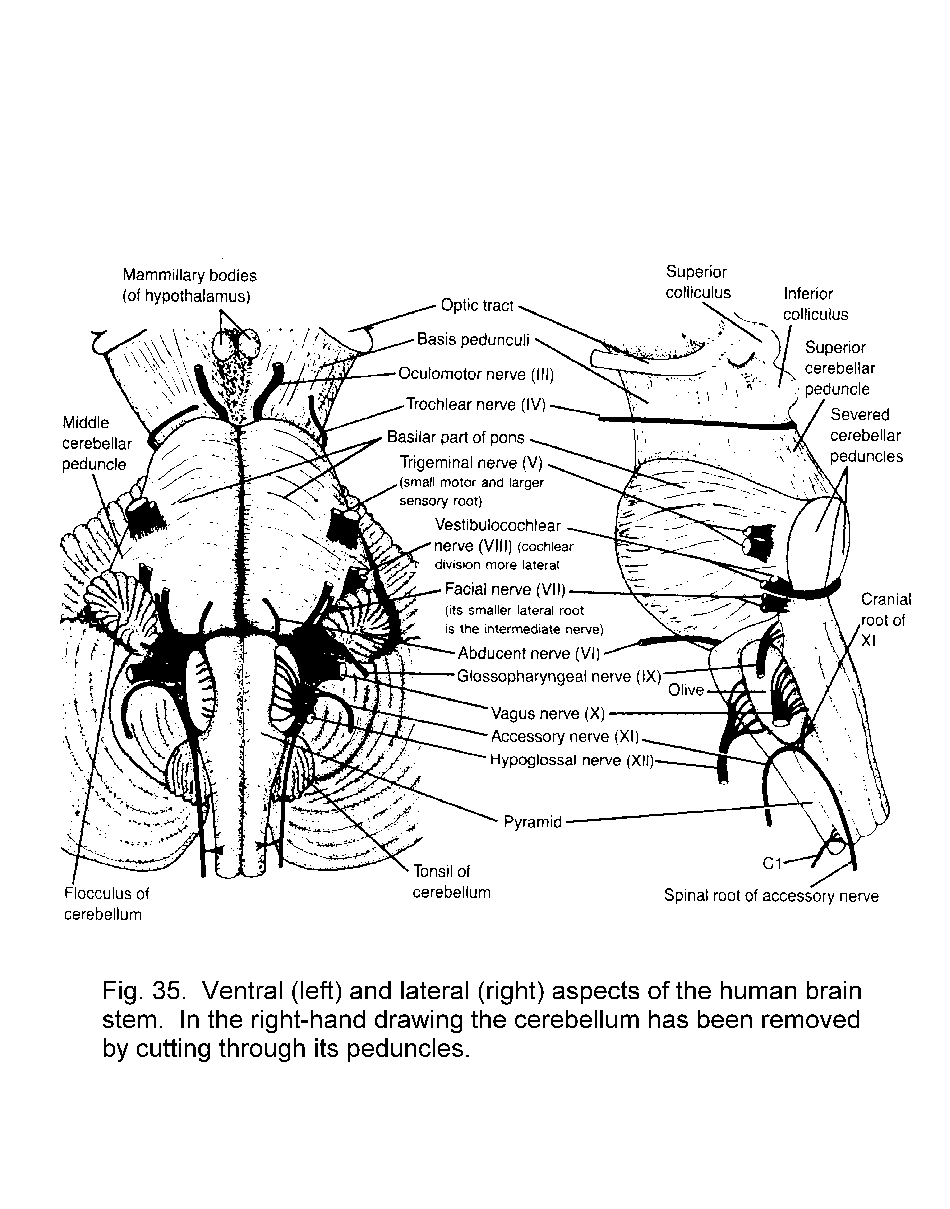{kind=link}
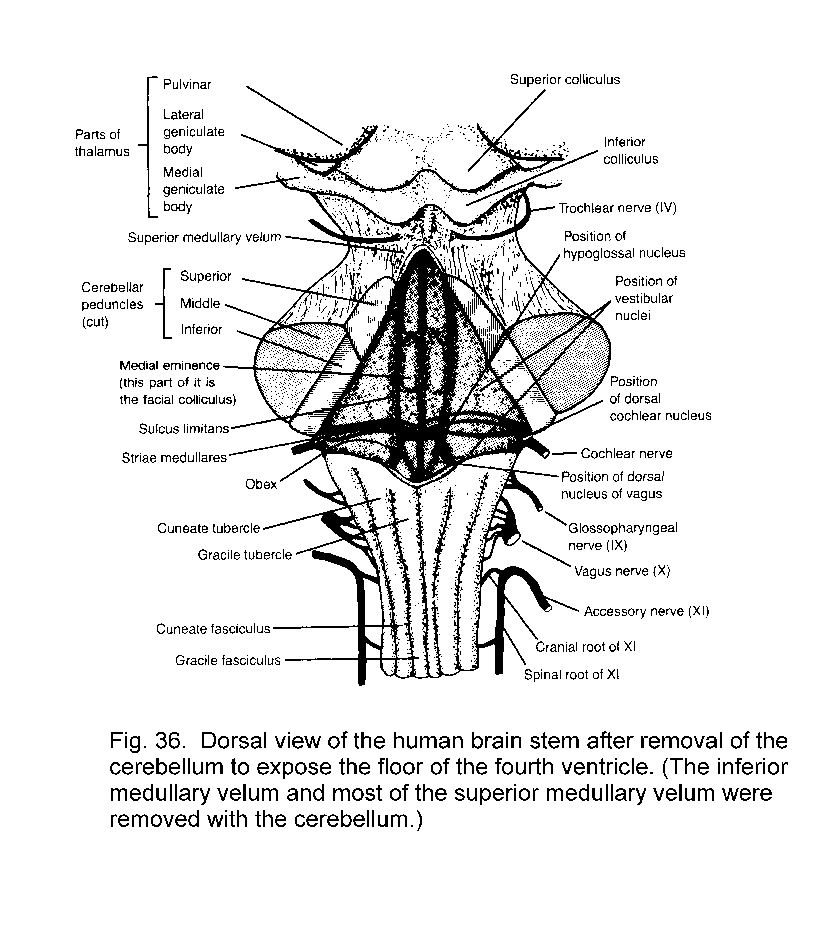{kind=link}
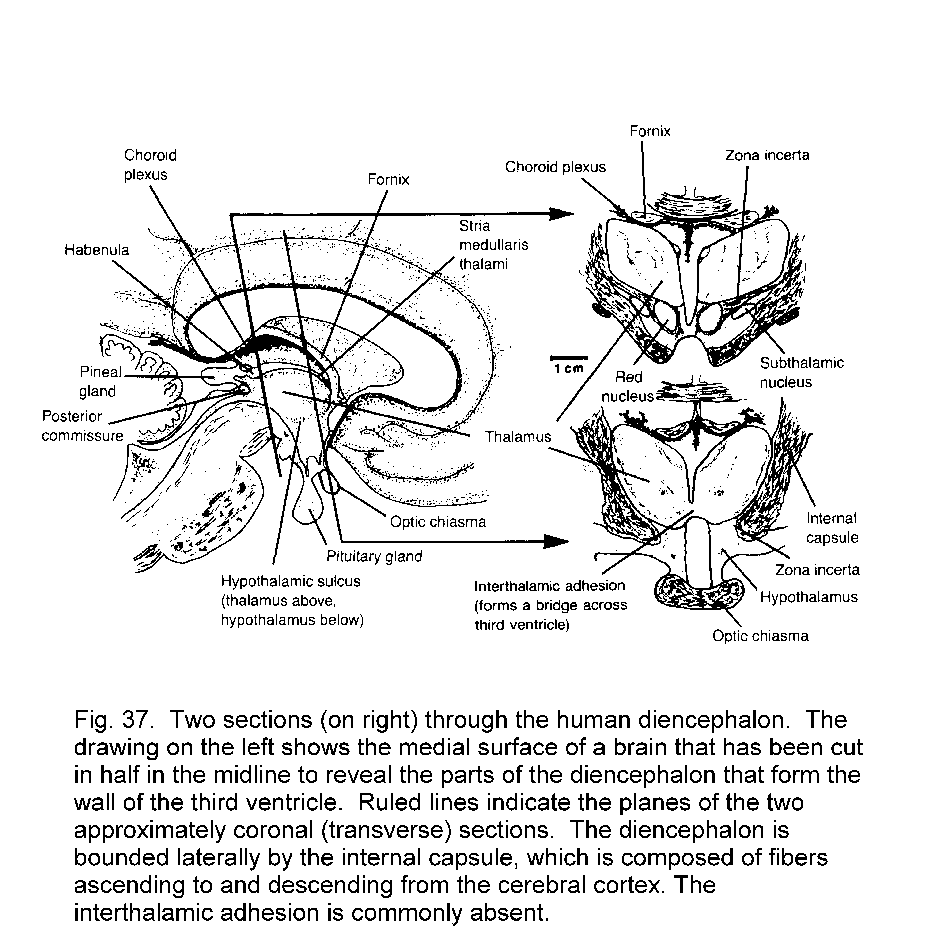{kind=link}
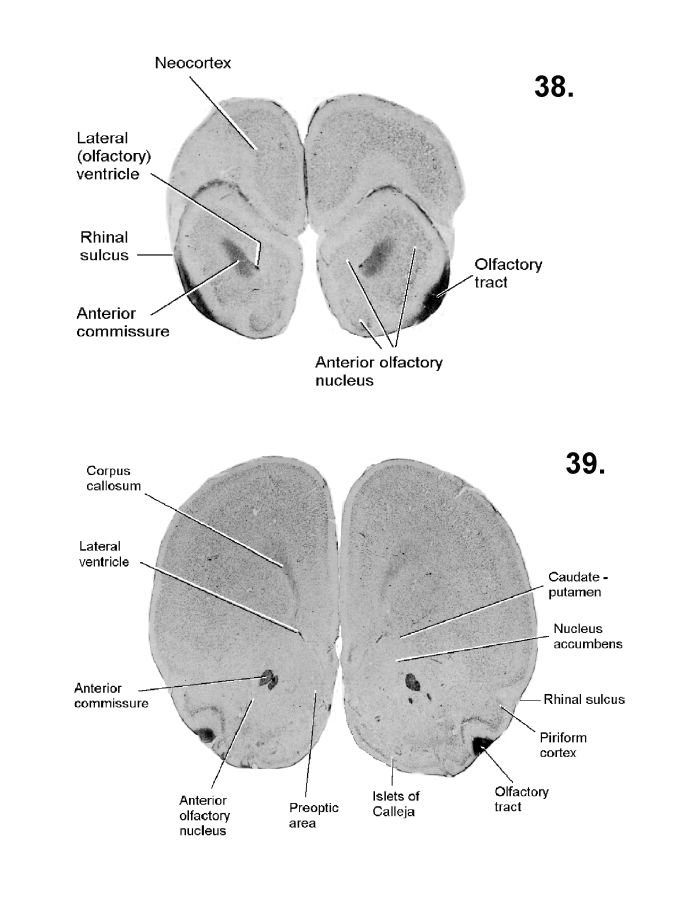{kind=link}
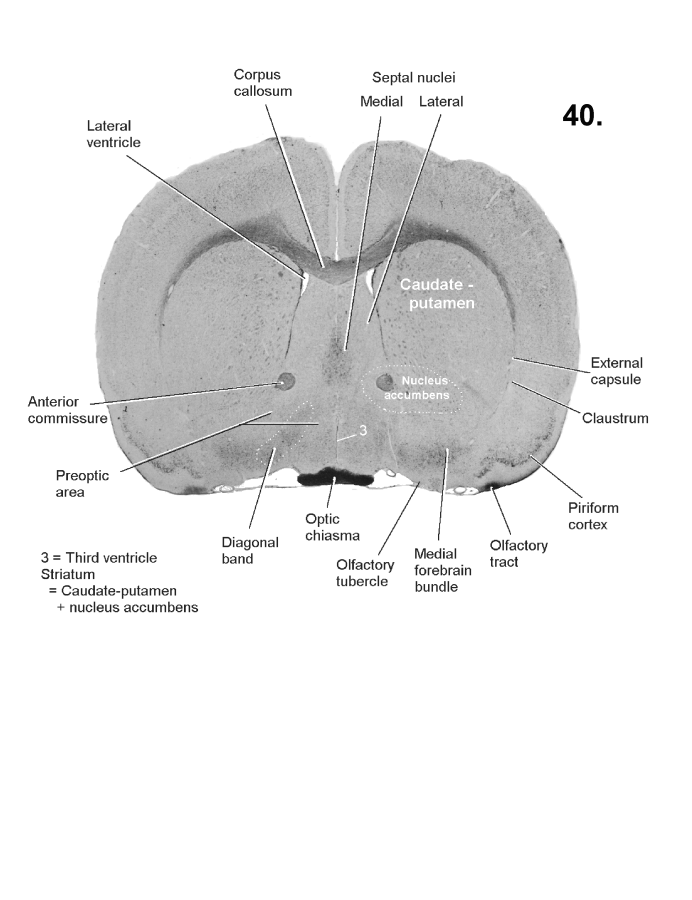{kind=link}
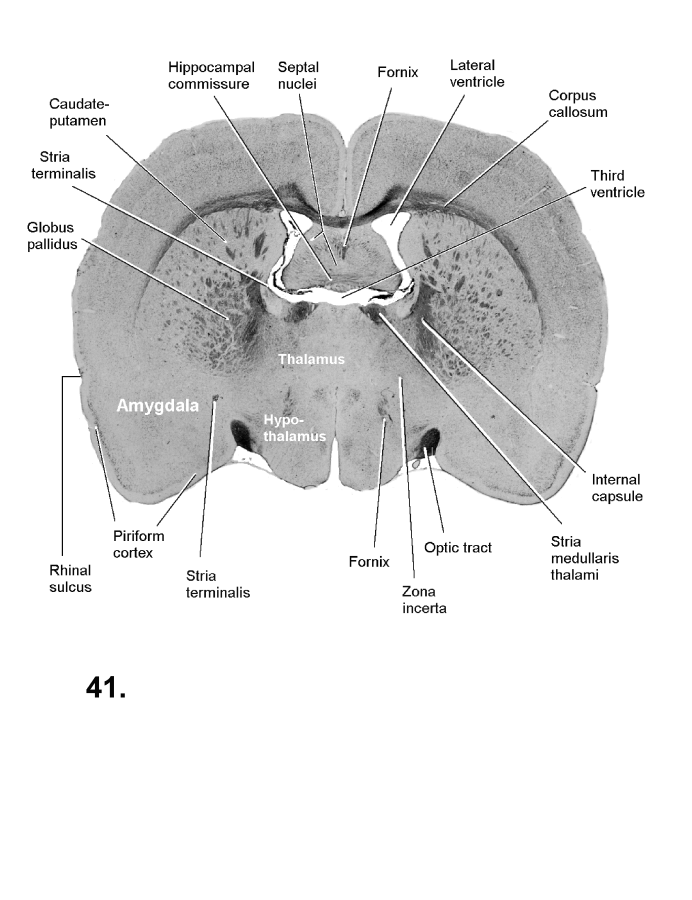{kind=link}
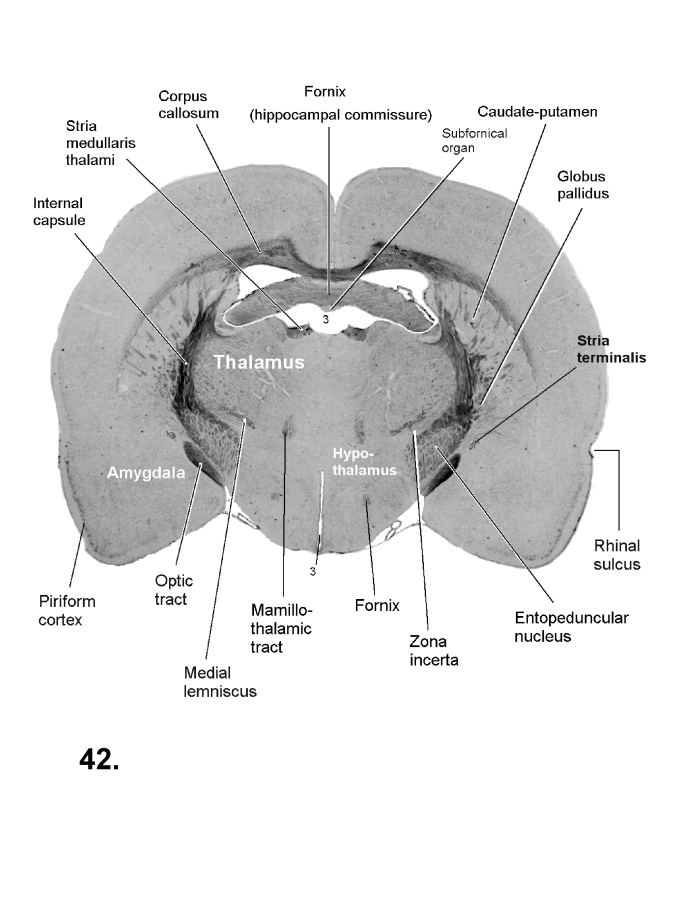{kind=link}
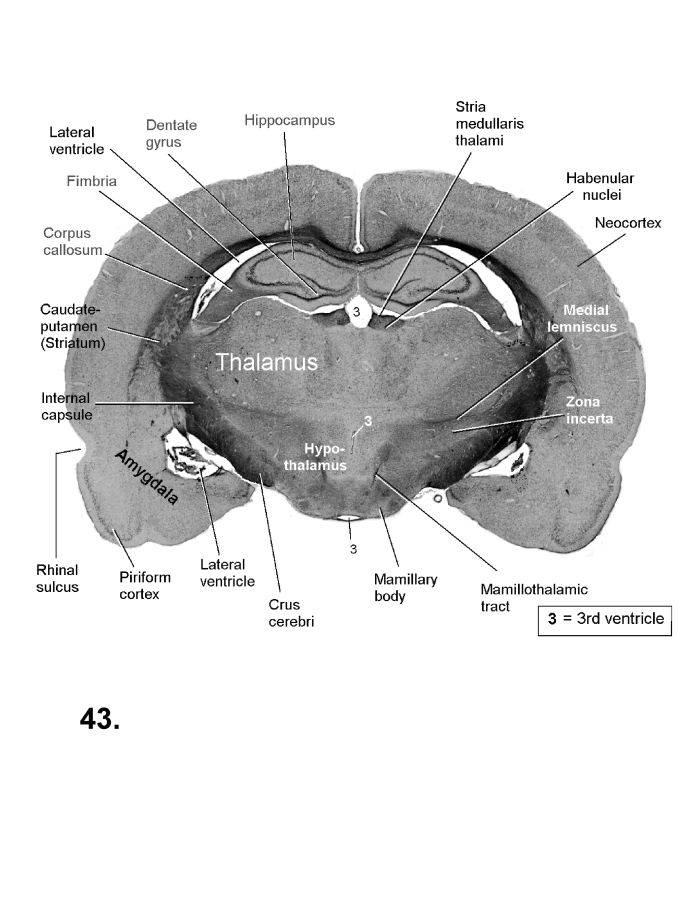{kind=link}
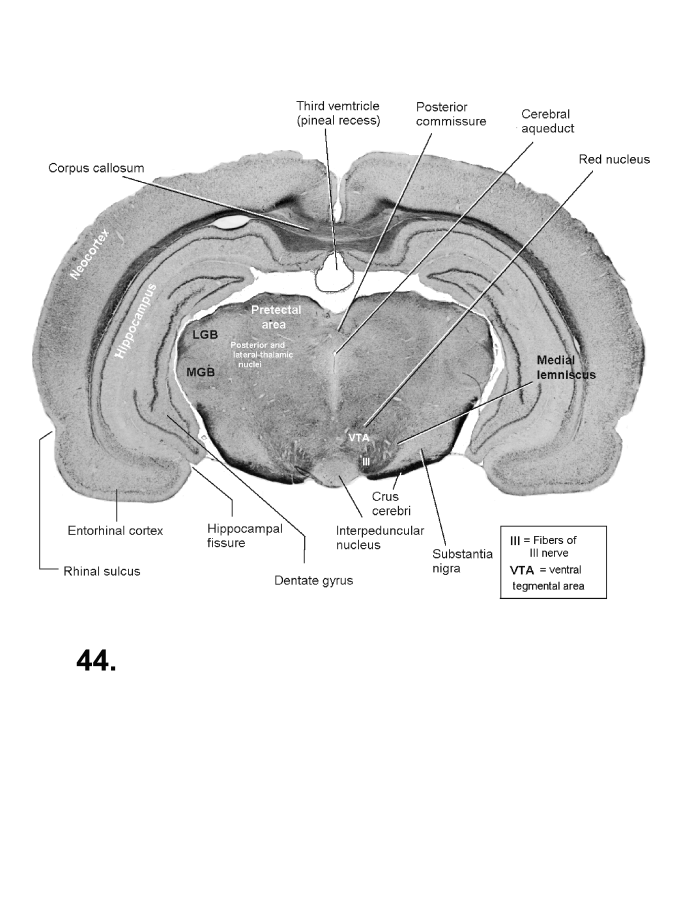{kind=link}
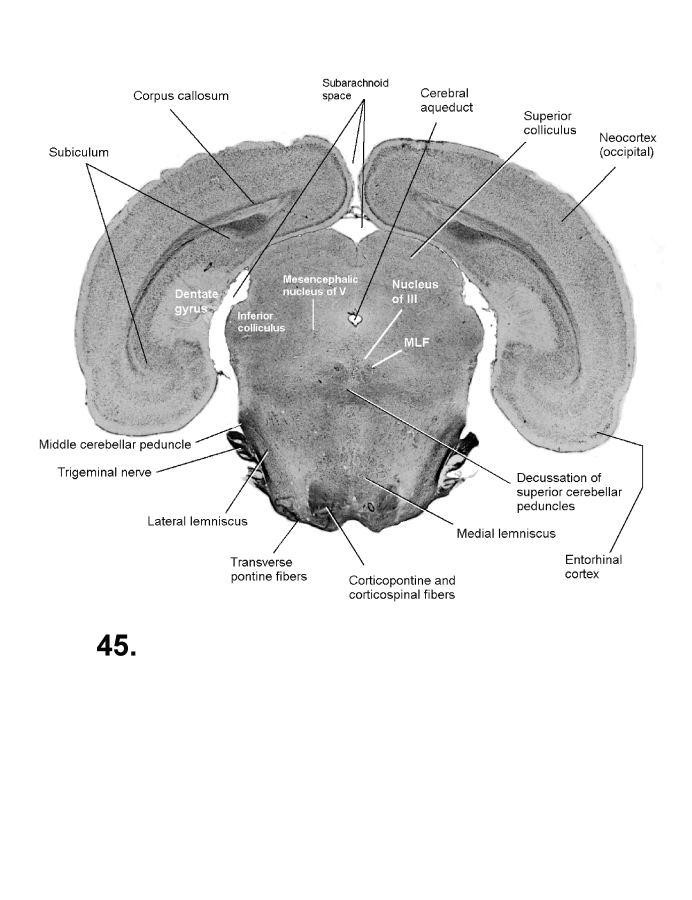{kind=link}
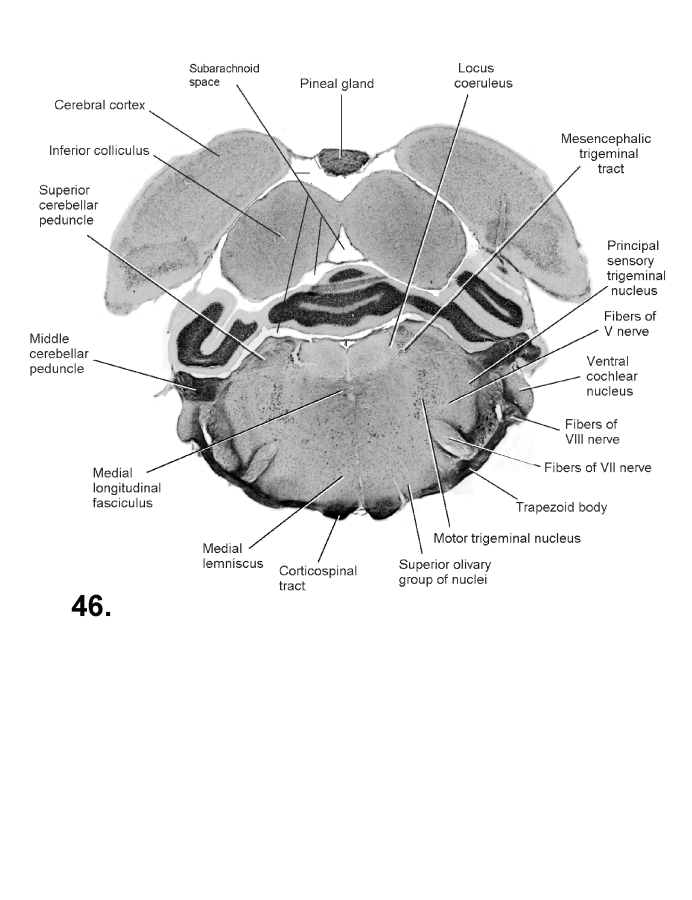{kind=link}
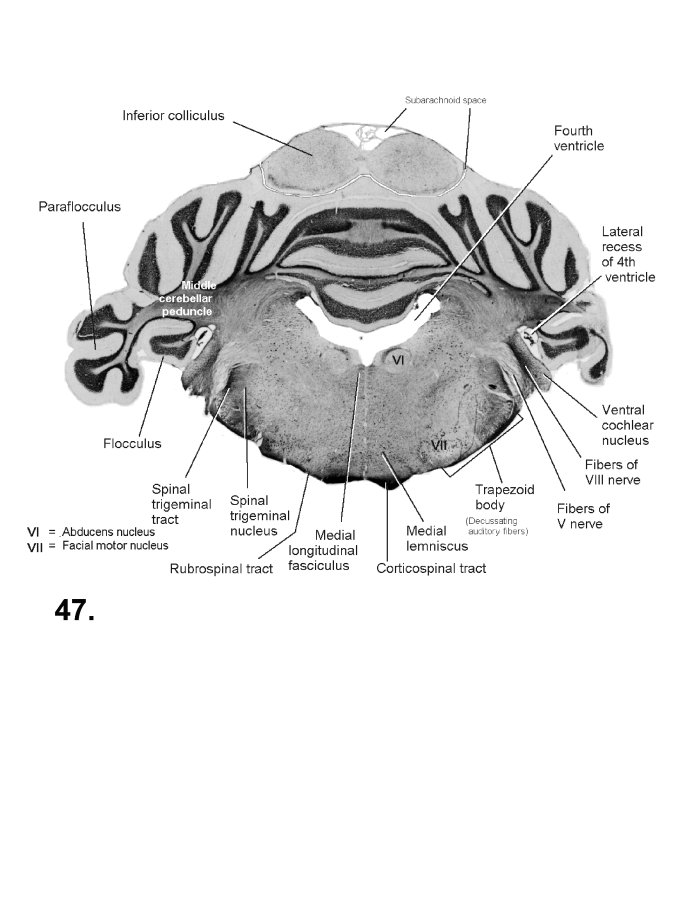{kind=link}
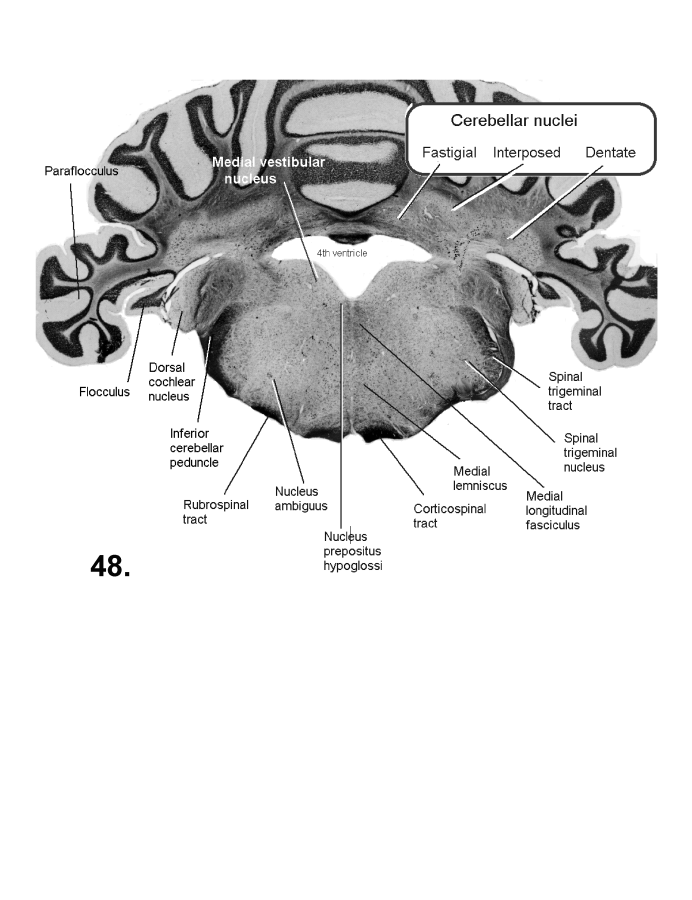{kind=link}
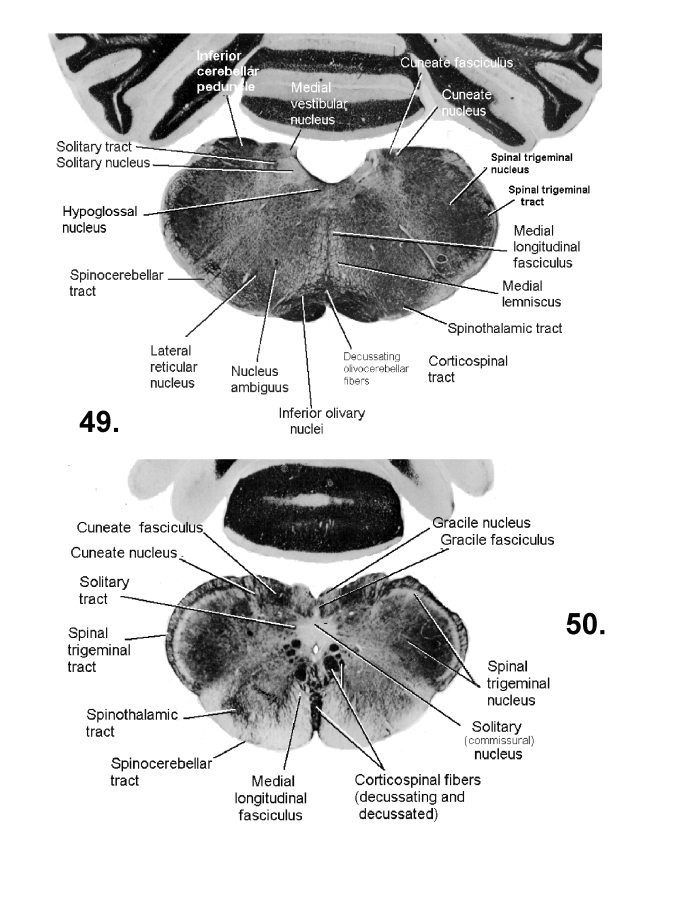{kind=link}
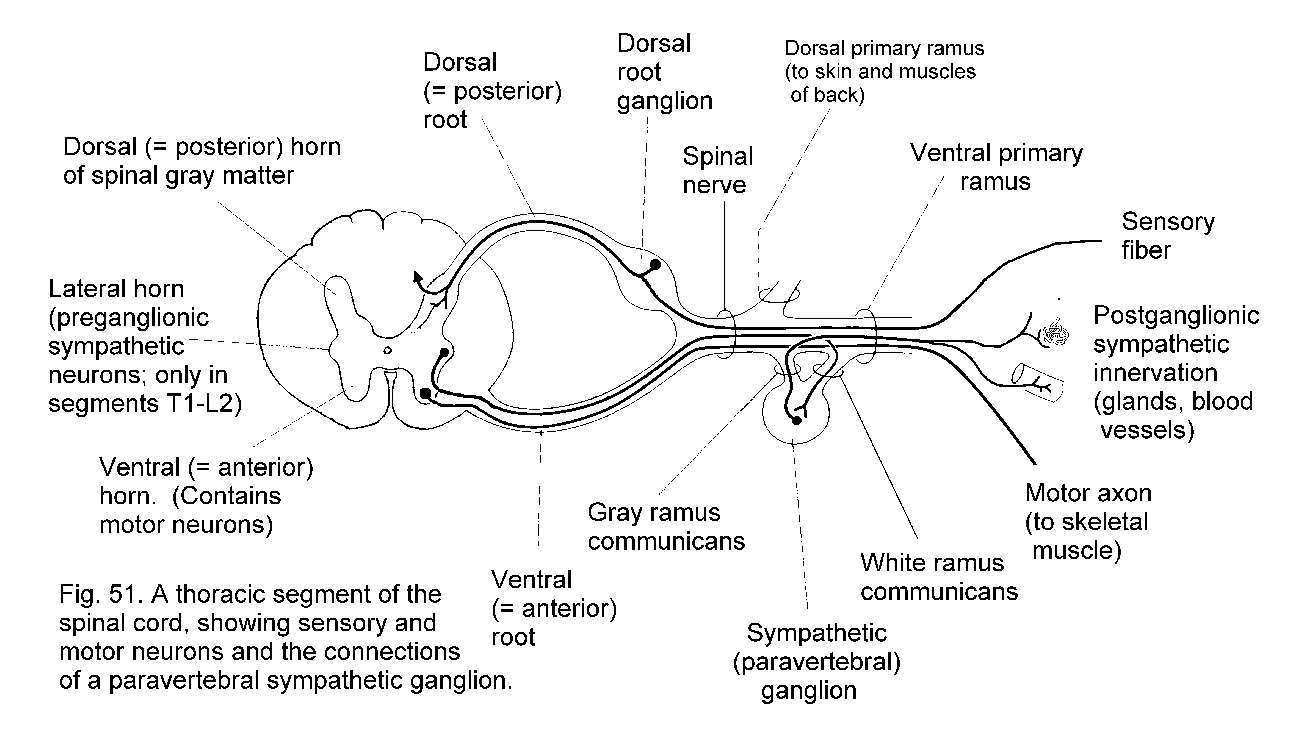{kind=link}
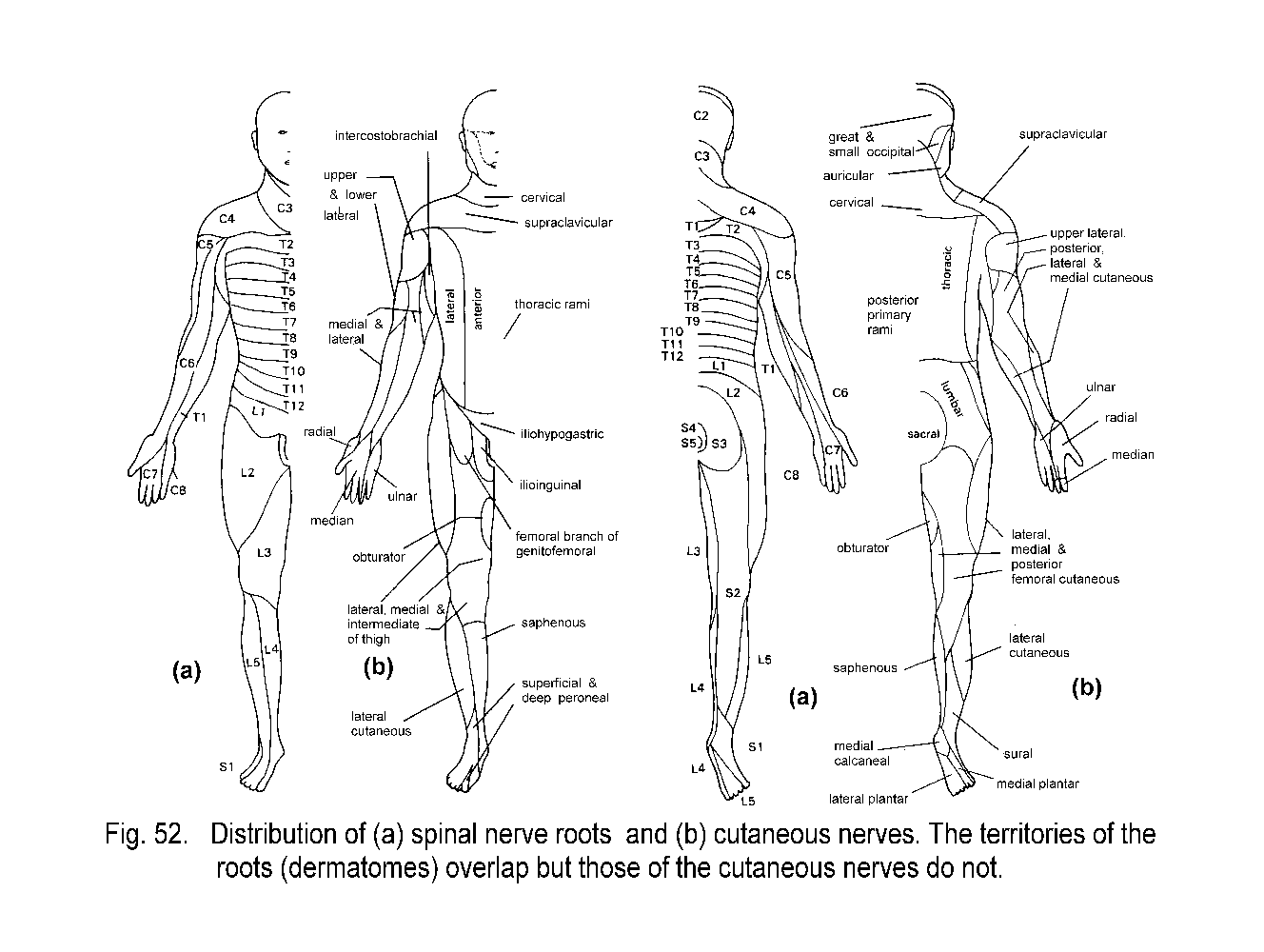{kind=link}
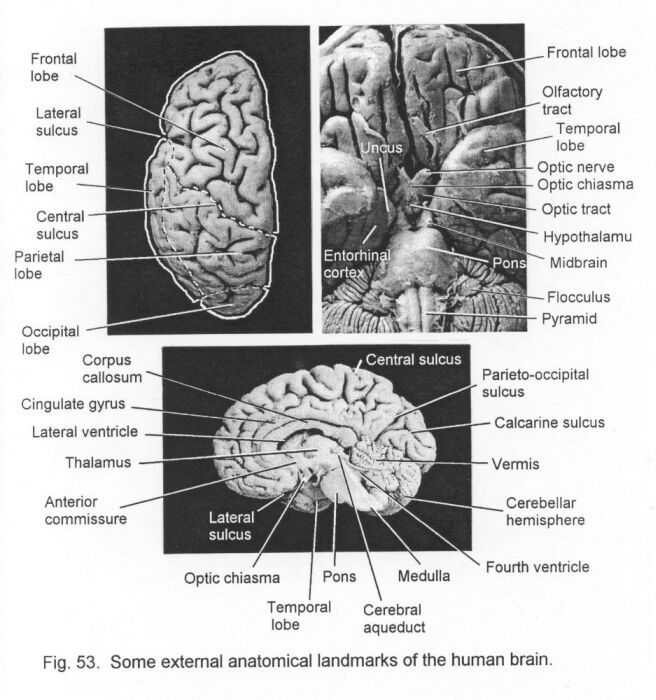{kind=link}
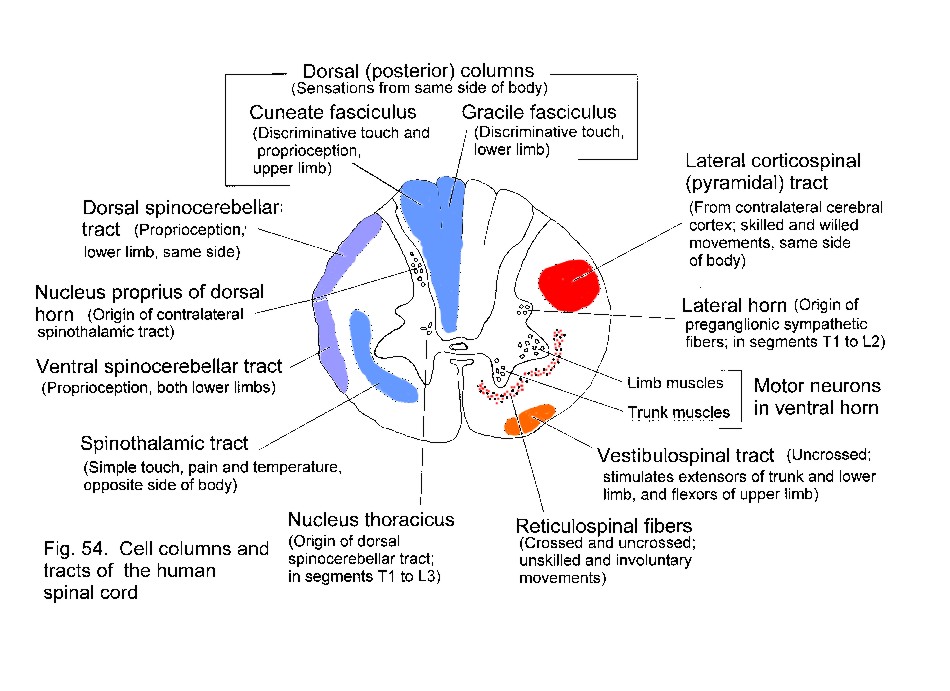{kind=link}
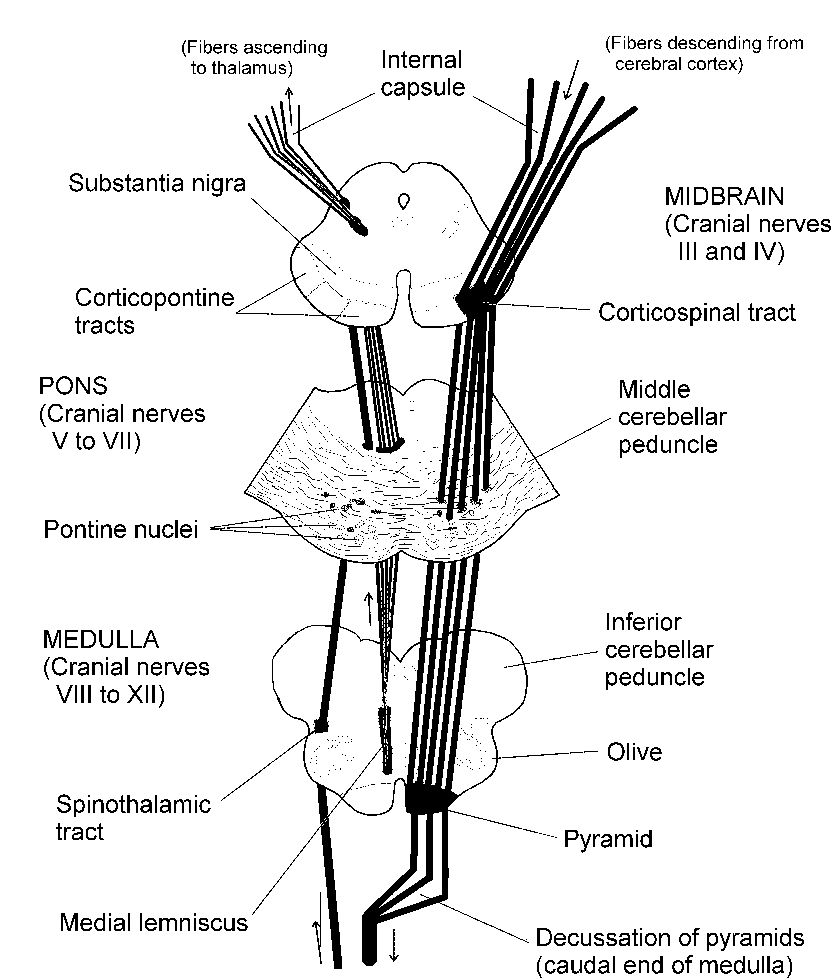{kind=link}
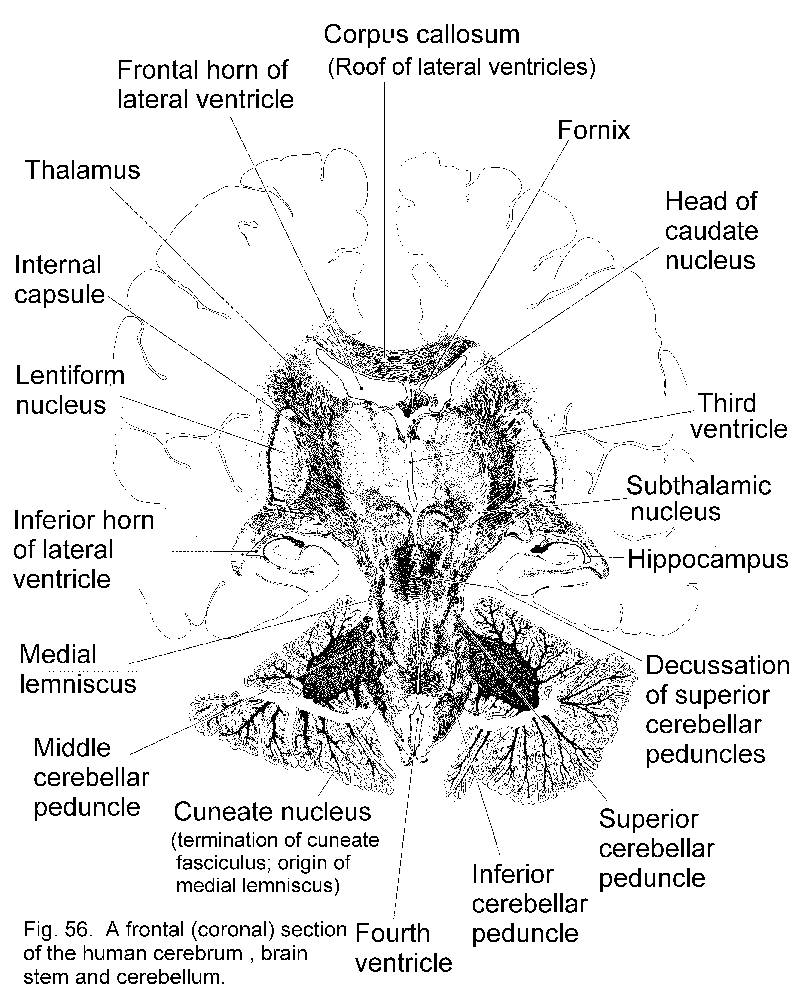{kind=link}
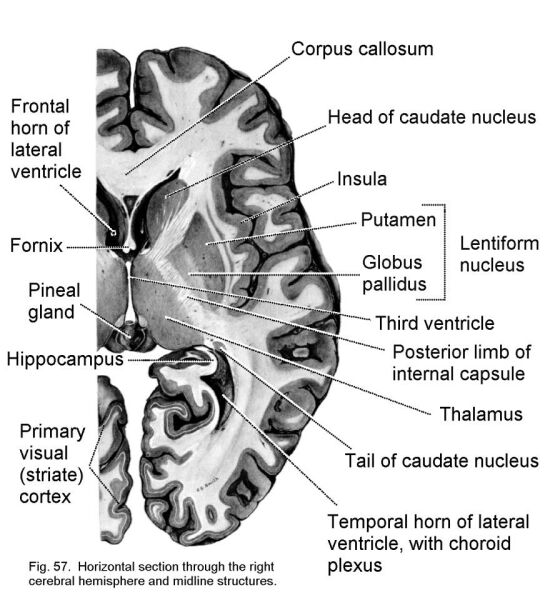{kind=link}
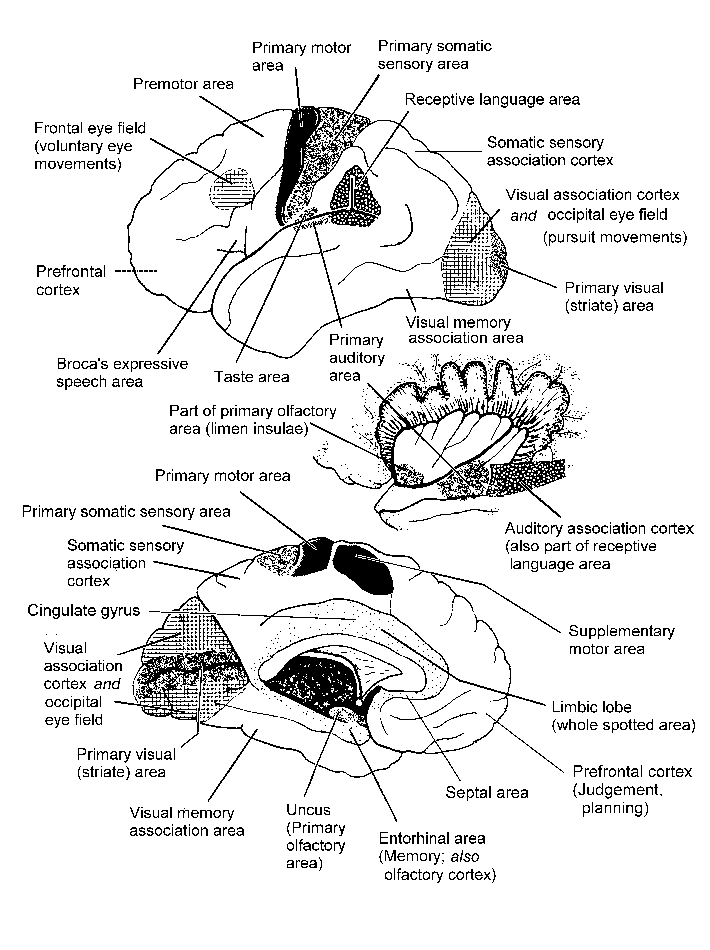{kind=link}
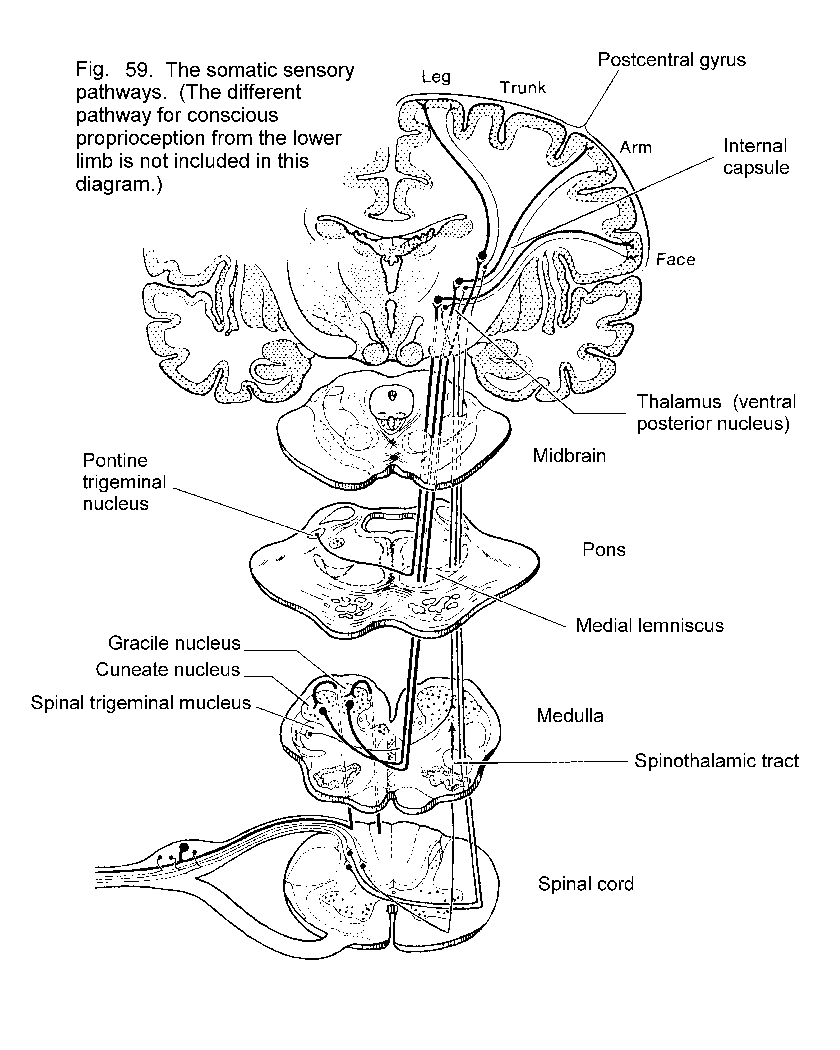{kind=link}
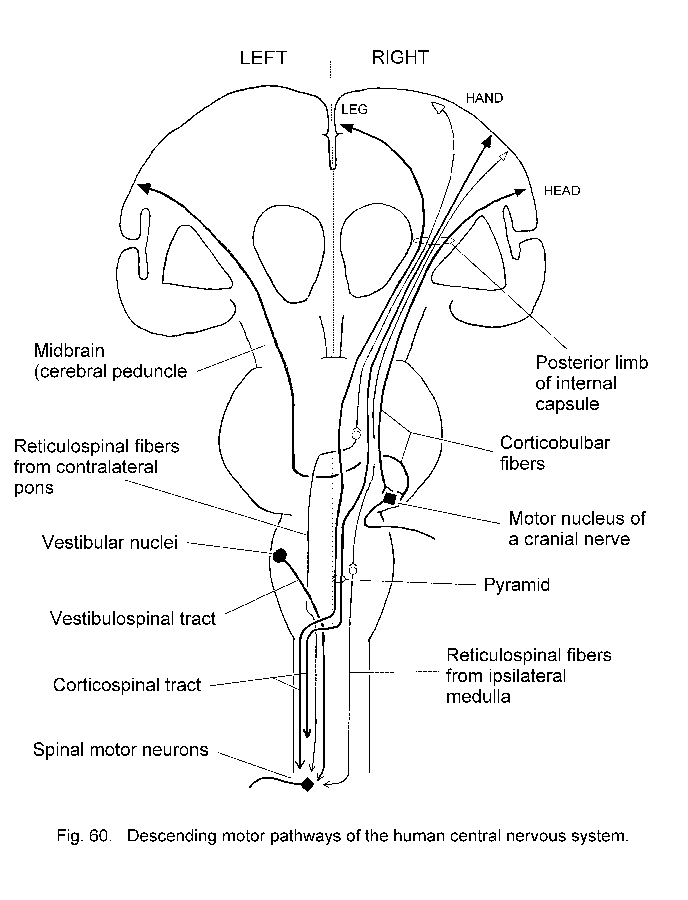{kind=link}
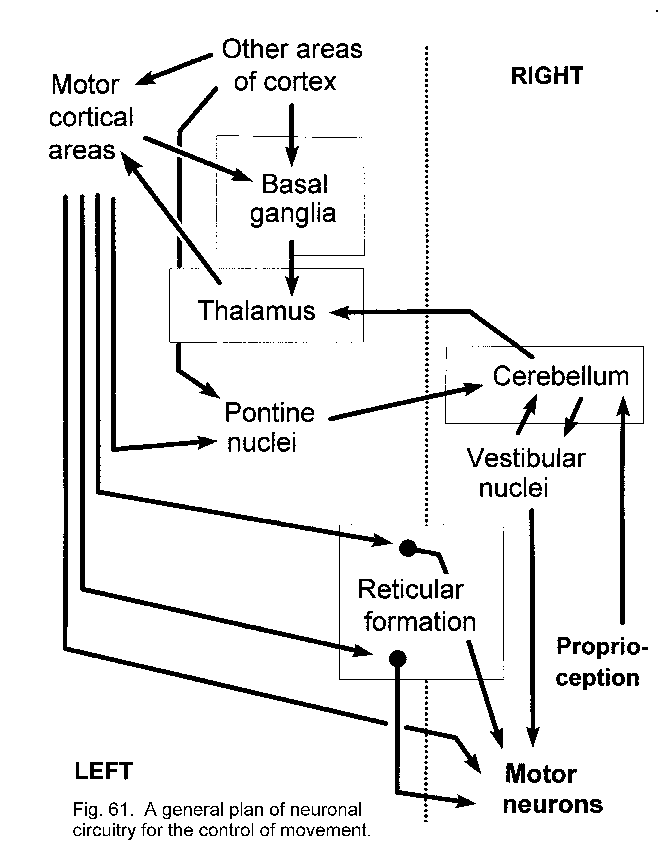{kind=link}
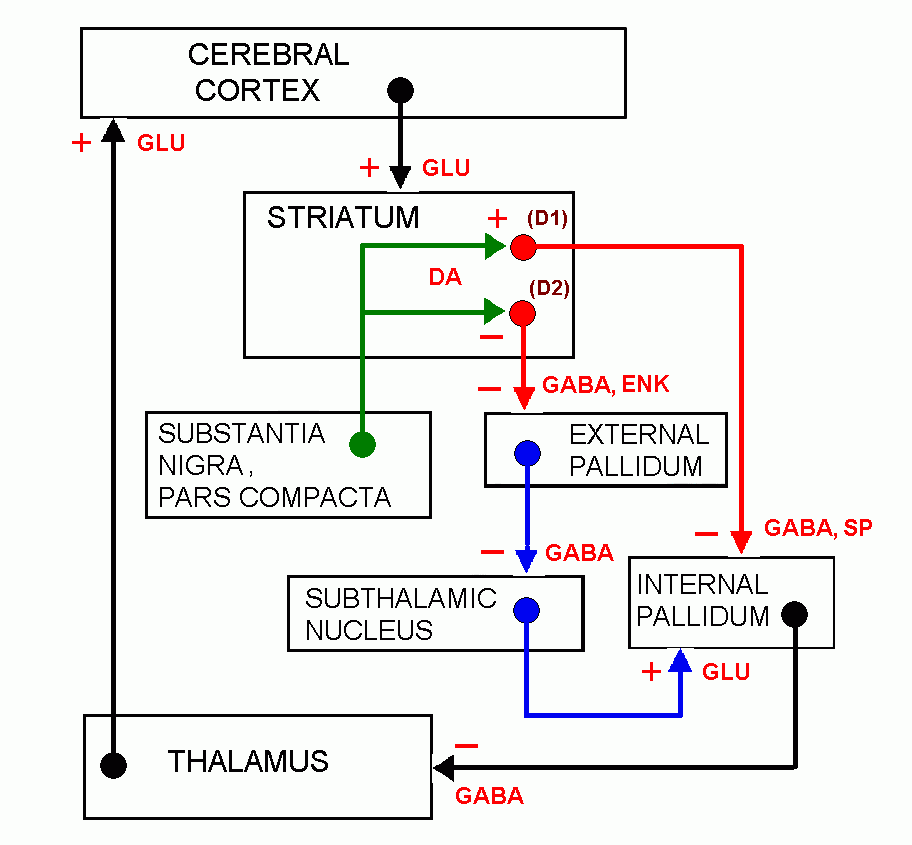{kind=link}
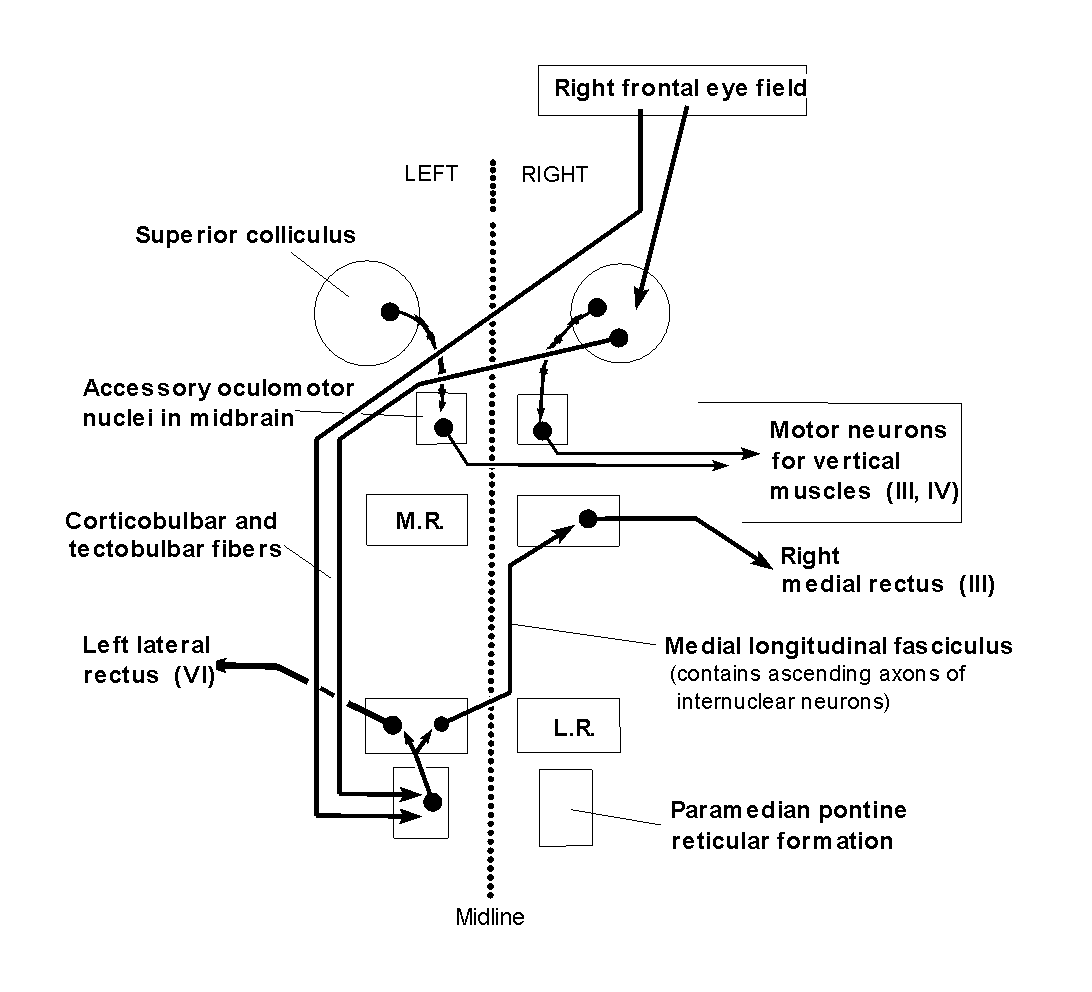{kind=link}
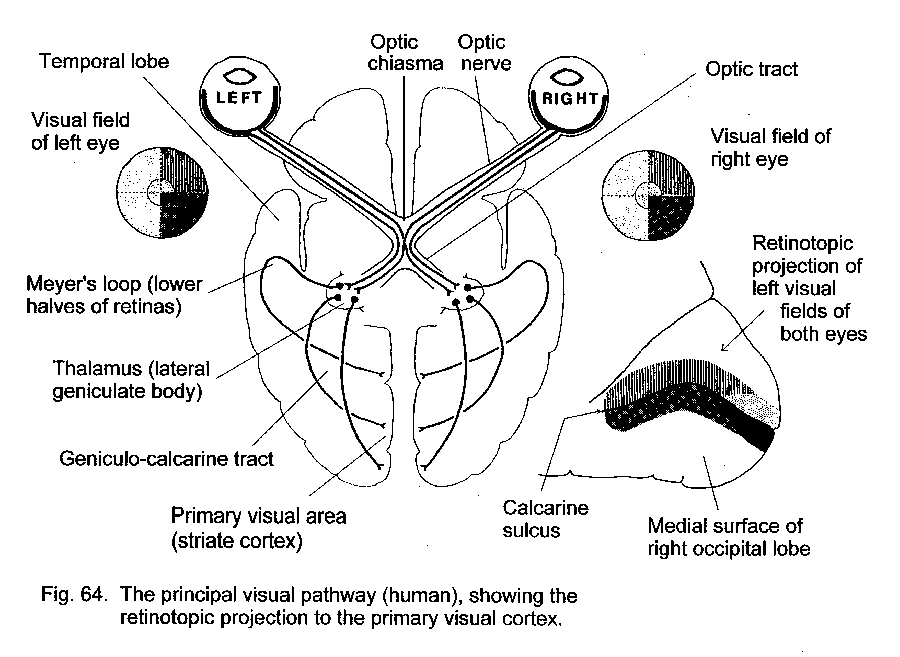{kind=link}