This is an excerpt from a much larger web document, Anatomical Foundations of Neuroscience, which is used as a resource by graduate students taking a
half course (Anatomy 535B) at the University of Western Ontario. Click on the link to view the complete document, which briefly reviews the anatomy of the nervous system in man and in other animals.
For Anatomy 350A (Functional Neuroanatomy) at UWO, this excerpt is a resource that compensates for the meagre treatment of the peripheral nervous system (and especially the autonomic system) in the recommended textbook.
As you read this document with your web browser, click on a highlighted link to look at an illustration (most are line drawings) or a table.
Use the browser's back button (such as Right-Click with Netscape) to
return to the text document.
You may save this document and any of its associated files and print them on paper for your own use. You may not distribute copies to other people. Although the facts in these files are common knowledge, the work needed to
present the information on a web page was done by me (J. A. Kiernan) in my employer's time.
This document is therefore Copyright (c) 2002 J A Kiernan and The University of Western Ontario.
This declaration means that nobody can legally copy the files for anything other than personal study.
Making copies to sell (or even give) to other students is forbidden and illegal.
Acknowledgement. I could not have put this together without some instruction in the arts of webbery, learned at one of Dr Candace Gibson's excellent faculty development courses.
J. A. Kiernan, Department of Anatomy and Cell Biology,
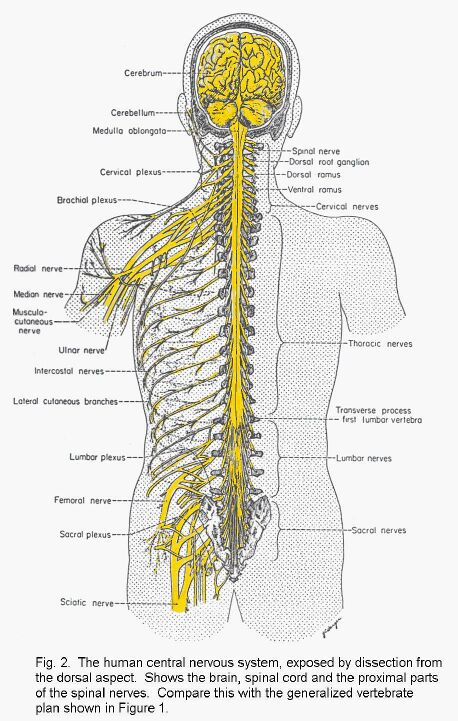
In man and in all other vertebrate animals, the nervous system has two divisions: the central nervous
system (CNS) is contained in the axial skeleton, and the peripheral nervous system (PNS) is distributed through most
of the other parts of the body (Fig. 1 - Plan of the human nervous system).
The central nervous system is connected to other parts of the body by nerves. Neuronal cell-bodies
outside the central nervous system occur in ganglia (singular, ganglion). The spinal nerves are
segmentally organized. Each has a dorsal and a ventral root, separately connected with the spinal cord.
The dorsal root bears a ganglion (called either a spinal ganglion or a dorsal root ganglion); the ventral
root does not. The dorsal roots are exclusively sensory in mammals, whereas the ventral roots contain
the axons of motor neurons and the axons of neurons that control internal organs, blood vessels, and
glands.
Paired cranial nerves connect the brain with other structures. The olfactory nerves, concerned with
smell, enter the olfactory bulb, which is at the rostral end of the telencephalon. The optic nerves, like the
retinas of the eyes, are made of central nervous tissue. They are therefore not real nerves but outgrowths
of the brain. The remaining cranial nerves emerge from the brain stem, which consists of the midbrain
pons and medulla.
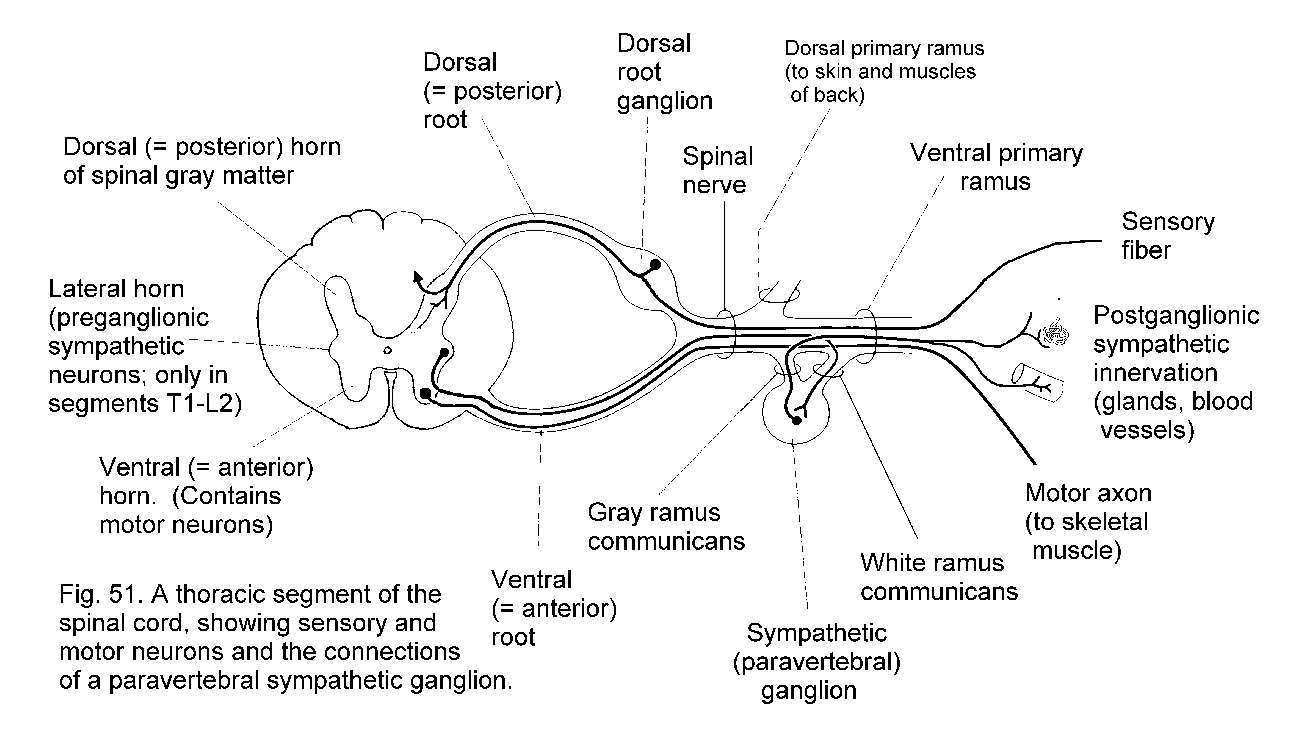
The nervous system develops from embryonic segments, but in the adult state this is obvious only in the connections of nerve roots with the spinal cord.
The formation of a spinal nerve is illustrated in Fig. 51 - Spinal nerve. This diagram also shows structural elements that will be referred to later in this Section. Spinal nerves have numbers derived
from the vertebrae. The highest spinal nerve penetrates the atlanto-occipital membrane, above the arch of the atlas, which is the first cervical vertebra or C1. The second cervical nerve passes between the atlas
(vertebra C1) and the axis (C2). There are 7 cervical vertebrae. The lowest cervical nerve is therefore C8. Cervical nerves 1 to 7 go through foramina above the numbered vertebrae. The roots of nerve C8 pass below
the arch of vertebra C7 and above that of T1. All the thoracic (T1 - T12), lumbar (L1 - L5) and sacral (S1 - S5) nerves go through foramina below the equivalently numbered vertebrae. To complete the story, a single
coccygeal nerve overlaps with S5 in supplying the perianal skin.
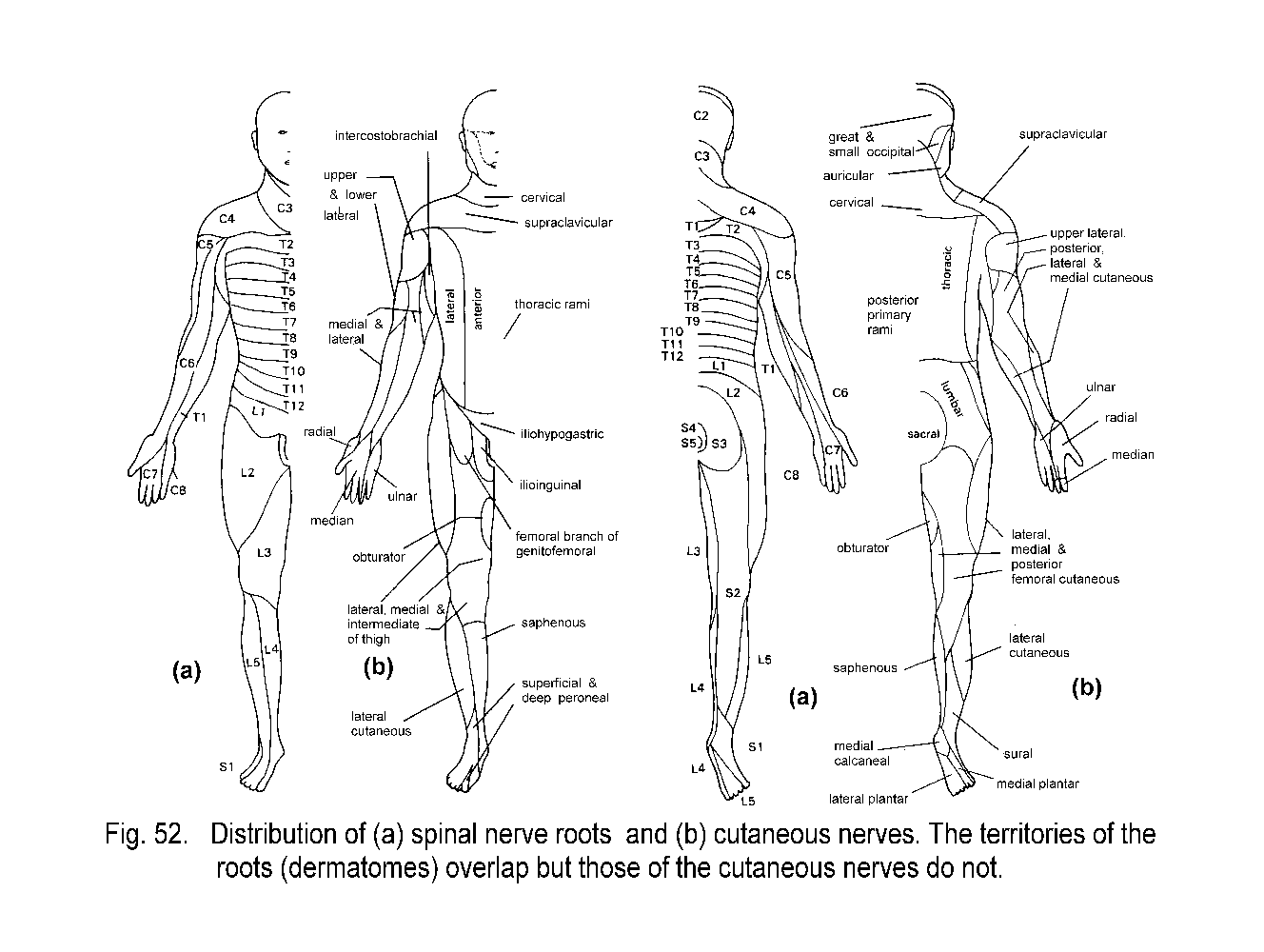
The most obvious consequence of the segmental organization of the spinal nerves is seen in the Dermatomes, which are bands of skin that run horizontally on the trunk and lengthwise on the limbs
(Fig. 52a - Dermatomes). Each dermatome is centered on the distribution of axons from a single dorsal root ganglion, but each ganglion also supplies skin in the dermatomes above and below its own level. Consequently, it
is necessary to transect three adjacent dorsal roots or spinal nerves in order to completely denervate the skin of one dermatome. Transection of a single spinal nerve, or destruction of its ganglion, diminishes but does not
abolish sensation in the affected segment of skin. The cutaneous lesions of herpes zoster, a common virus that infects certain pain-responsive neurons in individual sensory ganglia, often neatly map the distributions of
dermatomes and also illustrate the extension of innervation into the adjacent segments of skin. The nerve supply to the skin of the limbs is delivered by cutaneous nerves that are formed in limb plexuses
(brachial and lumbosacral) by complex interchanging and mixing of fibers from different spinal roots. The areas supplied by cutaneous nerves bear little resemblance to the dermatomes. They are sharply demarcated, with
little or no territorial overlapping (Fig. 52b - Dermatomes). The widely overlapping dermatomes cut across adjacent areas of skin supplied by cutaneous nerves. A cutaneous nerve lesion, such as an
injury or a mononeuropathy, results in a well defined area of defective sensation, and anatomical knowledge can be used to identify the affected nerve.
Most of the skin of the head is supplied by the three divisions of cranial nerve V. The areas are sharply demarcated, and therefore do not correspond to dermatomes. Cranial nerves VII, IX and X supply small, overlapping
areas of skin of the external ear, and the dermatome of the second cervical nerve includes parts of the head, ear, face and neck. (The first cervical nerve lacks a dorsal root in most people.)
Muscles receive motor and sensory innervation. Most of the muscles of the limbs are supplied nerves formed in the limb plexuses from two or more roots. Table 6 (Segmental landmarks) shows the
segmental innervation of a few clinically important muscles. A stretch reflex (tendon jerk) requires the integrity of both the motor and the proprioceptive sensory innervation of the muscle.
The vertebral column is longer than the spinal cord, which ends at the level of the upper border of vertebra L3 in the newborn and at the upper border of vertebra L2 in the adult. The lower spinal nerves must therefore
course caudally before passing through their corresponding intervertebral foramina. Immediately below the caudal end of the spinal cord, the neural canal contains the roots of nerves L2-L5, S1-S5 and the coccygeal nerve.
A lesion arising from the axial skeleton, such as a herniated intervertebral disk or tumor tissue from a vertebral body or pedicle, can press on the spinal cord or spinal nerves. The consequences depend on the level of the
involved disk or vertebra. In the cervical and upper thoracic spine there is little discrepancy between the spinal segments and the vertebrae. There is little free space in this part of the neural canal, so a lesion is
likely to impinge on the cord as well as on a spinal nerve. The body of vertebra T10 is level with spinal cord segment T11. Below this level, the discrepancy between vertebral and spinal levels increases rapidly, because
the lower lumbar and the sacral segments of the spinal cord are much shorter than the cervical and thoracic segments. All the spinal cord segments below T11 are in the range of just three vertebrae, T12, L1 and L2.
The foramina are above the levels of the intervertebral disks. Consequently, a herniated disk below C7 cannot compress its own segmental nerve; it presses on the nerve one or two segments lower.
For example, an L4-5 disk herniation commonly compresses spinal nerve L5 or S1, causing pain and other sensory abnormalities in the appropriate dermatomes see Fig. 52a).
Although the brain stem develops from segments (known as neuromeres), their peripheral distributions and central connections are most easily understood in terms of the functions of each nerve. These are set out in Table 7 - Cranial nerve components. Note that the second cranial "nerve," despite its traditional name, is not a nerve but an outgrowth of the brain, as is the retina.
Skeletal muscles are supplied by motor neurons whose cell bodies are in the spinal cord (anterior horn) or brain stem (motor nuclei of cranial nerves). In contrast, glands, cardiac muscle, and the smooth muscle of blood
vessels and internal organs are supplied by neurons in ganglia of the autonomic system. These ganglia receive afferent preganglionic fibers, which are the thinly myelinated axons of neurons in the spinal cord or
brain stem. The neurons in the ganglia have unmyelinated axons, the postganglionic fibers that innervate smooth and cardiac muscle and secretory cells. There are three divisions of the autonomic system: sympathetic,
parasympathetic and enteric.
The ganglia of the sympathetic system are the chains of paravertebral ganglia that lie on the lateral aspects of the bodies of the vertebrae, and also the preaortic or collateral ganglia
associated with the branches of the aorta that supply abdominal organs. There is a sympathetic chain ganglion for every spinal nerve. Postganglionic fibers enter the nerve by way of a gray ramus communicans (See Fig. 51 - Formation of spinal nerve) and are distributed to blood vessels, sweat glands and the little muscles that move hairs. Blood vessels of the skin constrict in response to their sympathetic
supply, whereas those in muscles dilate. Some of the ganglia for the nerves C1 to T1 are fused; consequently there are only three cervical sympathetic ganglia. In most people there is a stellate ganglion, formed from
inferior cervical (C7-C8) and first thoracic ganglia. The middle cervical ganglion is connected with nerves C5 and C6. Postganglionic fibers from the large superior cervical ganglion (C1-C4) accompany the
carotid artery and its branches. Some enter the eye, where they supply the dilator pupillae muscle of the iris. Others supply smooth muscle within the upper eyelid1. All three cervical ganglia send postganglionic fibers
into cardiac nerves, which run alongside the common carotid artery and aorta and supply the muscle of the heart. Increased activity of the sympathetic system increases the rate and force of contraction of the heart.
Peripheral nervous system
The University of Western Ontario, London, Canada
Structural plan of the nervous system
Segmental organization
Relation of spinal cord and nerve roots to the vertebral column
Cranial nerves
Autonomic nervous system
| In the absence of active sympathetic innervation of the eye, orbit and face, the pupil constricts (unopposed parasympathetic action), the upper eyelid droops partly, but can be raised voluntarily (intact oculomotor nerve innervation) and sweating does not occur on the affected side of the face. These changes (Horner's syndrome) can result from damage to the superior cervical ganglion, its pre- or postganglionic fibers, (which pass through the stellate ganglion), or to transection of descending axons in the lower brain stem or spinal cord that control the preganglionic neurons. |
Parasympathetic ganglia are found in the head, connected with certain cranial nerves, and associated with the walls of thoracic and pelvic viscera. Preganglionic fibers leave the brain stem in cranial nerves III, VII, IX and XI and terminate in cranial parasympathetic ganglia. The neurons in these ganglia supply the structures whose functions are stated in Table 7 - Cranial nerve components. The cardiac ganglia receive their preganglionic afferents from the vagus nerve; their neurons supply cardiac muscle cells, principally in the atria. The pelvic splanchnic nerves, branches of S2, S3 and S4, carry preganglionic fibers to the parasympathetic ganglia that supply the detrusor muscle of the urinary bladder and the blood vessels of erectile tissue in the genitalia.
The enteric nervous system consists of thousands of tiny, interconnected ganglia in the walls of the alimentary canal, from esophagus to anus, and of some of its associated structures such as the biliary system and pancreas. These ganglia, which supply the smooth muscle and secretory tissues of the gut, contain several types of neurons, with a wide variety of neurotransmitters. The enteric nervous system can do much of its work independently, but it is modulated by preganglionic fibers from the vagus nerve (to the stomach, small intestine and first half of the colon) and from the pelvic splanchnic nerves (distal colon and rectum). Parasympathetic activity stimulates propulsion of the contents of the gut. Of the vagal fibers that enter the abdomen, a majority end in enteric ganglia of the stomach, and the integrity of this preganglionic supply is essential for acid secretion and for opening of the pyloric sphincter.
Most of the postganglionic sympathetic fibers from the preaortic ganglia synapse with neurons in enteric ganglia, but some contact blood vessels and a few supply intestinal smooth muscle. Activity of the sympathetic system causes constriction of visceral blood vessels and retards propulsion of the contents of the alimentary canal.
--------------------------
Last updated: January 2003
{kind=link}
{kind=link}
{kind=link}