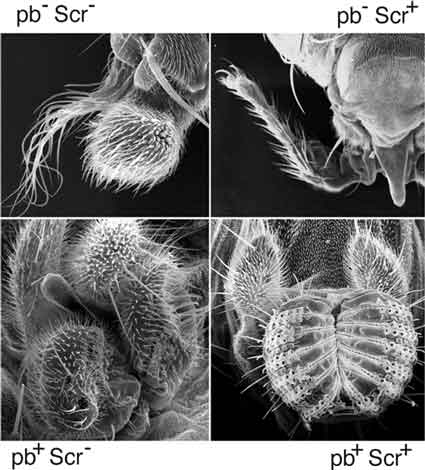
Role of PB and SCR in determining the identity of four body parts
What we have learned so far.
When I was a Graduate student studying sporulation in yeast, I was introduced to homeotic genes, and I was so intrigued by the phenotypes of legs sprouting from the head and extra pairs of wings that I decided to make these genes the focus of my future studies. In Drosophila there are eight homeotic genes clustered in two gene complexes. All the homeotic proteins contain a homeodomain. Interestingly the genes that control whether a yeast will sporulate or not, MAT
a2 and MATa1, are also homeodomain-containing proteins. Although it has been known for a long time that homeotic genes are required for the formation of particular body parts, it is still somewhat unknown how homeotic genes bring about the formation of particular body parts. Even knowing that homeotic genes encode a set of similar DNA-binding transcription factors has not lead to a good description of how a particular body part is formed apart from the general statement that the homeotic proteins regulate sets of genes that are required for the formation of body parts. It has only been in the past three or four years that this question has started to be addressed productively. I became interested in proboscipedia (pb) because it is the only Drosophila homeotic gene whose activity is not required during embryogenesis. I thought this adult viable property of pb null alleles made it a good candidate to address the problem of how a body part is put together. However, on the road to this long-term goal, I had to understand the phenotype of the pb null allele first. Why are there tarsi sticking out from the proboscis?
From the analysis of a series of genetic experiments, I was able to determine that the reason that pb mutants have tarsi sticking out of the proboscis is because Sex Combs Reduced (SCR) protein in the absence of PB protein directs tarsus identity (Percival-Smith et al., 1997). An experiment that shows this is when both PB and SCR proteins are removed, the proboscis is no longer tarsi but is now antennae. From this and other analyses, I was able to propose a model for how PB and SCR determine the identity four body parts: antennae, tarsi, maxillary palps and the proboscis. When PB and SCR are not expressed, antennal identity is determined. Expression of PB alone determines maxillary palp identity, and expression of SCR alone determines tarsal identity. Expression of PB and SCR together results in the formation of a PB-SCR containing multimeric protein complex that determines proboscis identity. Hence, proboscis identity is brought about by the combinatorial interaction between two homeodomain containing proteins PB and SCR, which brought me somewhat full circle because sporulation of yeast requires complex formation between the two homeodomain containing proteins MAT
a2 and MATa1.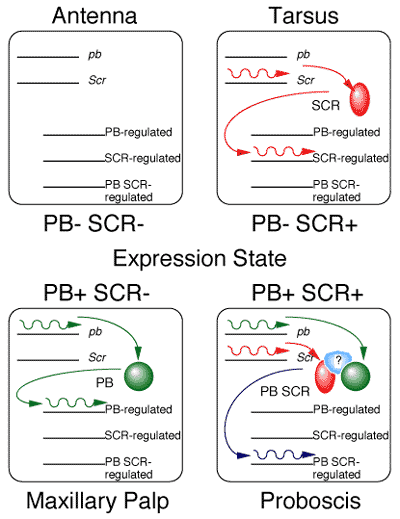
Where we are going.
In the analysis of the PB-SCR interaction, we found that it is spatially regulated. We propose that an additional factor(s) is required for PB-SCR complex formation. We have initiated a two hybrid screen to identify these additional factors. These factors are important for determining in which cell the PB-SCR interaction can occur.
We have also initiated a screen to identify PB-regulated genes using subtractive hybridization. We presently have a number of clones that encode transcripts whose transcription is PB-dependent. We hope these genes encode components important for the formation of the proboscis or maxillary palps.