In common terms, zoogeography is simply the study of geographical distribution of organisms about the globe.
It begins with the work of simple description: a compilation of the geographic ranges of species, which may then be collated into higher categories - genus, family, order and so on.
Explorers had always brought home traveler's tales of the strange organisms to be found elsewhere in the world, and, as exploration expanded and became scientific, such distributional information was seen as an important contribution to knowledge.
Such an enterprise was begun in earnest in the 19th. cent. and the founding compilation was that provided by Alfred Russell Wallace (1823-1913), co-discover (with Charles Darwin), of the process of Natural Selection.
He amassed much information first-hand from his explorations in South America and in South-East Asia (mainly what is now Indonesia and New Guinea), but the bulk came from a world-wide correspondence with other travelers and naturalists. The result was
and it still stands as a useful standard work in the
field.
His findings demonstrated that, in general, the different parts of the world supported greatly different assemblages of organisms - these findings provided ample confirmation of the impressions gained by generations of travelers that foreign parts were distinctive.
It was also abundantly clear that these geographical disparities in life were not haphazard or random, rather:
| This last point has recently been emphasised dramatically
by results from the analysis of DNA sequences from a diversity of African
mammal groups. Several of these animals have long been puzzling in
terms of their ancestry and relationships. The DNA data now shows
clearly that they form a single natural group now termed the Afrotheria.
This group includes such diverse forms as elephants, elephant shrews, tenrecs, golden moles, hyracoids, sirenians and aardvarks. The group makes very good geographic sense; the members are of African origin. Fossils and morphology had suggested that elephants, sirenians, and hyracoids, for example, are related to ungulates (horses, cows, etc.) but the new DNA evidence emphasises their peculiarly African status, distinct from these other lineages with many members found elsewhere in the world. |
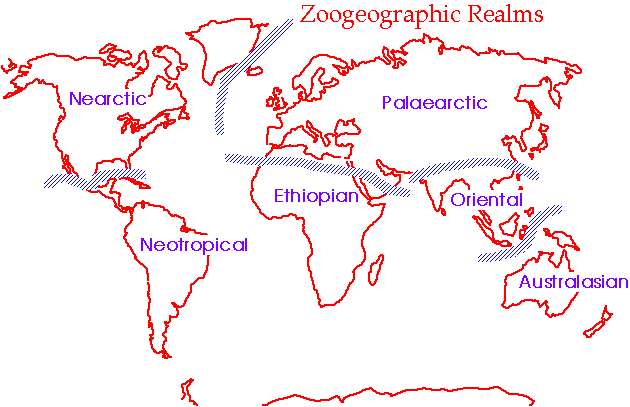
In other words, it became clear from the work of such as Wallace that continental regions were occupied by typical arrays of related organisms, different from other such regions. These regions (which didn't always exactly correspond to "continents") are termed Zoogeographic Realms (see below). There are similar regions defined on the distributions of plants.
Now why should this all be so? - after all, given the ruling idea of the times that things on earth were as decreed by the Deity, organisms could just as easily have been distributed in a diversity of other ways e.g. uniformly, or latitudinally, or according to where suitable habitat occurs (e.g. why were horses and cattle not found in the perfectly good grasslands of Australia or South America?).
This is the central question in biogeography, and it takes us from the realm of description into that of analysis.
This question was one of several other related questions about the nature of life on earth. For example, another big question was: why is there a natural hierarchical system of resemblance among organisms that allows us to easily classify them into unique, nested arrays - species within genera, within families, within orders.......... within kingdoms? Again, many other systems of resemblance are logically possible, so why this one?
The general grand answer to all these questions (which
themselves have a
very long history) was:
|
|
This, of course, is the process of evolutionary change and diversification, what Darwin in his great book On the Origin of Species called descent with modification, and it is the answer which we use to this day.
The deployment of this idea of the evolution of life against a historical backdrop of the earth's own changing face provides us with the answer to our central question: Thus we now see all of earth's organisms as each being part of an evolutionary tree of lineages, directly comparable to our own family lineages, though they reach vastly deeper into the past.
The branch points of this tree of evolutionary lineages - say the one separating marsupial and placental mammals, or the one separating those mammals which became the whales from those that became the hoofed mammals - are unique in time and space.
That is, lineages of organisms originate not only at particular points in evolutionary time but in particular regions of the globe.
These unique coincidences between lineage originations (branchings) and the particular parts of the earth where they happen have a profound impact on where the descendant organisms are found today.
Why is this so? It is so because different parts of the earth have had different histories of isolation from, and connection with, other parts of the earth,influencing where the lineage's constituent organisms may spread to.
In Darwin's day, although science understood the geological processes of erosion and sediment-deposition, and though it was clear that there had been dramatic changes in the elevation of parts of the earth's crust (Darwin found marine mollusc fossils thousands of feet up in the Andes mountains) it was supposed that earth's geography was static - that the map of earth had always been as it is today.
Thus in those days, distributions of organisms were interpreted in terms of today's geography. All this changed during this century.
Nowadays we understand that, far from being static,
the various crustal plates that make up the cores of continents have been
spinning and sliding about on the earth's surface throughout its enormously
long history, at times widely separated, at other times agglomerated
into large lumps.
| The web has a diversity of resources which make the complexities of ancient global geography more easy to comprehend. Here are two: 1. Global Earth History; 2. Earth History. |
We call this process continental drift. For example, today's Eurasian continent is composed of several core pieces that were once very widely scattered about the globe. And all this time, as land masses were skating about - now separating, now colliding - life's lineages have been evolving: very clearly, where you originate, and what happens to where you originated, have a profound impact of where you will eventually (today) be found.
Thus, some groups of organisms will have had initial access only to restricted regions of a single small crustal plate, while others will have had direct access to the wide-open spaces provided by several plates grouped into massive continents in which to spread and diversify. At later times, restricted lineages may have found their way to new continental regions as plates joined up - say when ground sloths spread into North America as it joined to South America. Other lineages, having spread over a continent made of several plates, may then have had their range greatly split up as the plates drifted apart. This happened when the ancestors of the ratite (flightless) birds, which originated in the ancient super-continent of Gondwanaland, became widely separated as South America, Antarctica and Australia drifted off to their present far-flung locations uring the Mesozoic. Ther they evolved into their present-day forms: ostrich, rheas, emu, cassowaries, kiwis and moas. Such massively disjunct distributions as that of the ratite birds provided major (though fruitful) problems for Darwin, Wallace, and other early biologists in their attempts to explain patterns of distribution: How on earth could they get from one place to the other, flightless as they were, when separated by thousands of kilometres of open ocean? But they didn't know about continental drift.
We may also look at a smaller scale example a little closer
to home: humans originated in Africa, and eventually walked to Asia
and Europe. For hundreds of thousands of years they had been restricted
to parts of the world with dry-land connections between them. But
when, during the last ice-ages, a broad land bridge was formed between
Asia and Alaska, they could walk on and colonise the whole New World.
| SIDE-NOTE:
As well as helping us understand the distribution of present-day organisms,
continental drift theory has permitted some dramatic predictionsabout
where on the earth we may expect to find fossils of particular kinds of
creatures. For example, our knowledge that South America, Antarctica
and Australia were joined together until the mid-Cretaceous (around 100
million years ago) suggested that we ought to be able to find fossils,
in the currently lifeless Antarctic, of creatures known to have
been widely distributed on earth in those times. This expectation
has been fulfilled, and several reptiles are now known from Paleozoic and
Mesozoic Antarctic deposits.
Similarly, though marsupial (pouched) mammals are found in both South America and Australia, monotremes, the egg-laying mammals, are today known only from the Australian region. This was always a puzzle - they ought to have occurred in South America too, at least in the past; this puzzle was solved recently when fossil monotremes were found in Cretaceous deposits in Argentina. Perhaps we shall eventually find monotreme fossils in Antarctica........... |
and the key realisation is that these barriers have been temporary and mobile: just as the continents have shown a great many different conformations, so the world's oceans have shifted, opened and closed - they have been enormously variable in their location and extent. Thus for example:
|
Continental drift and the unique time-sequence of lineage origination, then, are what explain the existence of what we call the zoogeographical realms. |
As was thoroughly catalogued by Wallace, as one travels from continent to continent, there are often massive shifts in the presence and/or abundance of organismal groups. That is, some groups show substantial continental endemism - they are restricted to certain continental regions. As defined by the distribution of animal groups, there are six realms:
Regions defined on plant distributions are somewhat different. Of course, organisms can, and do, spread out from their evolutionary origin to reach new and distant regions even if their continents don't join up, but this spread is limited by a) the dispersal capacities of the organisms themselves (some can fly, others must walk etc.), b) by the extent and kinds of obstacles in their way (e.g. rivers, oceans, deserts, mountains), c) by time, and d) by climate.
Further, organisms can go extinct from parts of their range, even that of their origin, e.g. camels & horses, though they evolved N. America, went extinct there.
Given the enormous spans of time since many lineages evolved, it is perhaps amazing that many aspects of distribution still bear the marks of ancient lineage originations and continents; other aspects reflect more recent events, including recent climate.
Examples of endemic mammal families of biogeographic realms:
A more precise notion of the degree to which the different regions resemble and differ from one another is to tabulate the numbers of, say, families unique to, or shared by, the various regions. Below is a table showing the pattern of occurence of the 79 families of terrestrial mammals, which are a good group to consider, since they have rather limited dispersal capacities (so their original distributions won't get quickly blurred). For this reason, the several families of bats are excluded altogether because, being capable of flight, they have very different powers of dispersal from the other mammals. Also excluded are eleven families which have successfully dispersed to all but one or two regions - the Soricidae (shrews), Sciuridae (squirrels), Cricetidae (voles, hamsters), Muridae (rats, mice), Leporidae (rabbits), Cervidae (deer), Ursidae (bears), Canidae (dogs), Felidae (cats), Mustelidae (weasels, skunks), and Bovidae (cattle, sheep, goats, antelope etc.) To have included these families would be to obscure the patterns. The numbers on the diagonal are those restricted (endemic) to a given region; the off-diagonal numbers refer to those shared by any two regions.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
So what are the patterns? Well, first be sure to understand the table. Look at the first column: we can see that 4 families are unique to the Nearctic, while it shares 6, 5, 0, 1, and 0 with the Neotropical, Palæarctic, African, Oriental and Australian realms respectively. Families shared by more than two realms will be counted in more than one cell, so totals are not always possible to figure out - for example, one of the six Neotropical families shared with the Nearctic realm is also shared with the Palæarctic, and so that family gets counted twice.
But, to the patterns. First note that 51 (4+15+1+16+5+10) families (73%) are endemic to one realm or another. Next note that all Australian mammal families are endemic (zero shared with other regions), and that the Neotropical and African realms are also high in familial endemism, with 15 and 16 families respectively. However, the percent endemism of each of these two region is quite different, because of their different totals: 68% for the Neotropical and 52% for the African realms. Finally, note that the Palæarctic region shows very low endemism (~5%).
Patterns such as this are clearly a reflection of the duration and degree of isolation of the different continental regions during the time of rapid mammal evolutionary radiation (late Mesozoic/early Cenozoic - the time when Pangaea broke up, and broad ocean barriers made Africa, Australia and South America island continents for millions of years).
In the next class will look at some details of these patterns:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}